
Промышленное скрещивание животных
Промышленное скрещивание преследует пользовательные (производство мяса, молока, яиц, шерсти, рабочая производительность) цели. Применяется в товарных хозяйствах. Промышленное скрещивание имеет две формы — простое и сложное.

Пример простого скрещивания
Простое скрещивание
Простое cкрещивание — это скрещивание двух или нескольких пород для получения помесей первого поколения или второго поколения животных, используемых для производства продукции и исключения их дальнейшего разведения.
Этот метод разведения широко практикуется в скотоводстве, свиноводстве, птицеводстве практически во всех странах мира с развитым животноводством. Основная задача — получение помесных гетерозиготных животных с большой продуктивностью. При промышленном скрещивании используется специфическая комбинационная способность, приводящая к эффекту гетерозиса.
Сложное скрещивание
Сложным (ротационным, переменным) скрещиванием называется такое скрещивание, при котором попеременно чистопородных маток исходных пород, а затем и помесных подбирают к производителям исходных пород. В данном случае только для получения первого помесного поколения необходимы чистопородные матки одной из используемых в скрещиваниях пород: в последующих же поколениях чистопородными производителями исходных пород осеменяют полученных помесных маток.
Видообразование
Ч. Дарвин считал, что видообразование включает дивергенцию внутри вида, вымирание промежуточных форм и выживание крайних как проявление конкуренции.
Согласно современным взглядам, видообразование – это процесс перехода генетически открытой системы популяции в генетически изолированную. Основными формами видообразования являются географическое и экологическое.
Географическое (аллопатрическое) видообразование основано на пространственной изоляции, которая в дальнейшем сопровождается экологическими и поведенческими изменениями, приводящими к биологической изоляции. Это довольно медленный процесс формирования и обособления новых видов за счет обособления географических рас, ареалы которых не перекрываются ареалом материнского вида. Сначала географические расы превращаются в подвиды, затем в полувиды и далее – в новые виды. Чаще всего географическое видообразование связано с расчленением ареала и разделением широко распространенного вида на популяции с полной изоляцией друг от друга. В качестве примера можно привести галапагосских вьюрков, впервые описанных Ч. Дарвином. Образование разных видов вьюрков происходило на разных островах и шло по пути аллопатрического видообразования.
Экологическое (симпатрическое) видообразование базируется на экологической изоляции. Прежде всего, этот способ связан с приспособлением популяции к определенной пище, тейпературе, глубине, т. е. к определенным условиям среды. При этом формирующийся новый вид находится в пределах одного ареала с материнским видом. Репродуктивная изоляция может формироваться и поддерживаться благодаря разобщению сезонов размножения. Примером может служить возникновение ранне- и поздноцветущих рас у погремка.
Влияние отбора на количественные признаки
Основное влияние отбора заключается в изменении частоты проявления генов в популяции, которая в то же время при естественном отборе всегда связана с адаптивной ценностью признака или группы признаков. Поскольку количественные признаки обусловлены большим числом генов и находятся под сильным влиянием факторов окружающей среды, действие отбора на изменение частот генов выражается через изменение фенотипической ценности как средних значений данного признака в популяции, их вариансы и ковариансы. Существуют три основные формы воздействия естественного отбора на количественные признаки в популяции: стабилизирующее, направляющее и разделяющее.
Стабилизирующий отбор. Стабилизирующий отбор (часто его называют также оптимальным, сбалансированным или отбором на среднее значение признака) наблюдается у особей со средним значением выраженности количественного признака, обладающего самой высокой адаптивной ценностью. В потомстве таких особей среднее значение признака не изменяется или же из-за утраты в ряду распределения одинаковой части крайних вариантов уменьшается его варианса.
Направляющий отбор. Вследствие отбора адаптивная ценность может возрастать или снижаться вместе со средним значением данного количественного признака. Направляющий отбор приводит к увеличению выраженности среднего значения признака в последующих поколениях, при этом варианса может измениться или же остаться постоянной.
Разделяющий отбор. Если в одной и той же популяции крайние варианты обладают лучшей адаптивной ценностью, чем варианты, имеющие обычно приближенно среднее значение адаптивной ценности, то это приводит к разделяющему отбору на плюс- и минус-варианты. При этом среднее значение адаптивной ценности может оставаться неизменным в последующих поколениях и тем не менее привести к увеличению значения вариансы.
Во всех перечисленных случаях успех отбора зависит от того, в какой мере фенотип является отражением генотипа, точнее от характера наследуемости данного количественного признака.
Отборы, проводимые в селекции растений, в большинстве своем относятся к направляющему типу, при котором особи с лучшей средней ценностью выбираются в качестве родительских форм следующего поколения. Отбор по некоторым экономически важным признакам часто бывает ограничен из-за слабой жизнеспособности чрезмерно отселектированных фенотипов как следствия стабилизирующего естественного отбора, препятствующего действию искусственного отбора. В естественных популяциях разделяющий отбор способствует проявлению генетического полиморфизма, в искусственно созданных популяциях его применяют редко.
Синтетическая теория эволюции
В современной науке существует несколько эволюционных теорий, которые обладают рядом отличий. Ключевой из них является объяснение механизмов эволюционных процессов. Наибольшее признание завоевала синтетическая теория эволюции. В ее основе лежит дарвиновское учение, развивающееся с помощью актуальных научных достижений.
Начало синтетической теории отмечено 40-ми годами XX столетия. Данная теория является синтезом разных предметных областей научных знаний, таких как дарвинизм, генетика, молекулярная биология. Большой вклад в развитие синтетической теории внесла наука палеонтология.
Данный подход способствовал созданию более полной картины развития жизни на Земле. Согласно синтетической теории эволюции, дарвиновское учение не опровергнуто, а, наоборот, дополнено. Многие тезисы, которые выдвинул ученый, получили подтверждение в настоящее время, так как в тот период, когда они были сформулированы, отсутствовали необходимые ресурсы для их доказательства, что было связано с недостаточным уровнем развития научной мысли.
Понятие «синтетическая» было сформулировано и активно употреблялось, благодаря одному из создателей синтетической теории эволюции Дж. Хаксли. В 1942 году ученый опубликовал труд под названием «Эволюция – современный синтез». Стоит отметить его большую роль в формировании Всемирного фонда дикой природы. Дж. Хаксли привнес огромный вклад в развитие науки, за что был удостоен медалью Дарвина и рыцарского звания.
Благодаря синтетической теории эволюции, значительно расширяются представления Дарвина об эволюционных процессах за счет раскрытия разнообразных сил эволюции, которые являлись триггерами. В учениях Дарвина упоминается лишь внутривидовая и межвидовая борьба за выживание при воздействии неблагоприятных факторов. В современной науке к факторам эволюции относят мутацию, изоляцию, популяционные всплески численности и другие.
Синтетическая теория основана на двух теориях:
- микроэволюция, изучающая изменения на генетическом уровне, за счет которых образуются новые популяции;
- макроэволюция, исследующая закономерности и направления эволюции живых организмов, в том числе, происхождение человека.
Макроэволюция и микроэволюция являются процессами, которые протекают под влиянием природных изменений. События, которые происходят на микроэволюционном уровне, являются доступными для наблюдений. Процессы макроэволюции занимают тысячелетия и могут быть только реконструированы.
В таком случае, отсутствуют какие-либо возможности для постановки эксперимента, свидетельствующего о том, что макроэволюция существует. С другой стороны, ученым удалось найти косвенные доказательства в процессе изучения и развития наук:
- палеонтологии;
- морфологии;
- археологии;
- генетики.
Основные положения синтетической теории эволюции в упрощенном виде:
- эволюция происходит за счет наследственных изменений, таких как генные мутации;
- главным фактором эволюции является естественный отбор как итог борьбы за существование;
- наименьшая единица эволюции представляет собой популяцию;
- целость вида сохраняется с помощью миграций особей между популяциями;
- новый вид образуется благодаря длительному эволюционному процессу;
- эволюция обладает направленным характером, т. е. не имеет определенности ее конечной цели.
Вид, его критерии и структура
Одно из первых определений вида предложено К. Линнеем: вид – это совокупность особей, обладающих общими морфологическими признаками и не скрещивающихся с другими группами.
Современное определение вида: вид – это совокупность географически и экологически близких популяций, способных в природных условиях скрещиваться между собой, обладающих общими морфофизиологическими признаками, биологически изолированных от популяций других видов.
Вид является генетически изолированной системой.Признаки, отличающие один вид от другого, называются критериями вида:
- Морфологический критерий – это характеристика особенностей строения вида. С его помощью легко определить виды, не являющиеся близкими, однако необходимо учитывать, что многие морфологические признаки оказываются характерными не только для конкретного вида, но и для рода, семейства или отряда. Также верно и обратное утверждение: сильные морфологические отличия между особями не всегда говорят об их принадлежности к разным видам (породы собак, сорта растений).
- Генетический критерий – каждый вид имеет свой кариотип, характеризующийся определенным числом хромосом, их структурой и дифференциальной окраской. Использование этого критерия позволяет эффективно различать даже близкородственные виды.
- Географический критерий включает ареал обитания вида. Для видов-космополитов этот критерий утрачивает свое значение.
- Экологический критерий описывает непосредственное местообитание вида – его экологическую нишу, которая у каждого вида своя (это понятие включает в себя особенности питания, предпочтительные места обитания и размножения и т.д.). Однако разные виды могут занимать сходные экологические ниши.
- Физиологический критерий основывается на отличиях между видами в протекании процессов (активность ферментов, синтез и накопление различных веществ). Каждый из критериев имеет свои недостатки и преимущества, но по отдельности они часто недостаточны для различения видов. Для точного определения вида необходимо применение совокупности критериев.
Фиксация генов
Дисперсные процессы в бесконечно малой популяции могут действовать до определенных границ, так как частота проявления гена может находиться только в пределах от 0 до 1. Так, когда ген достигает частоты 0 или 1, дальнейшее его изменение возможно лишь при мутации. Аллель, достигший частоты 1, фиксируется в популяции, достигший 0 — устраняется
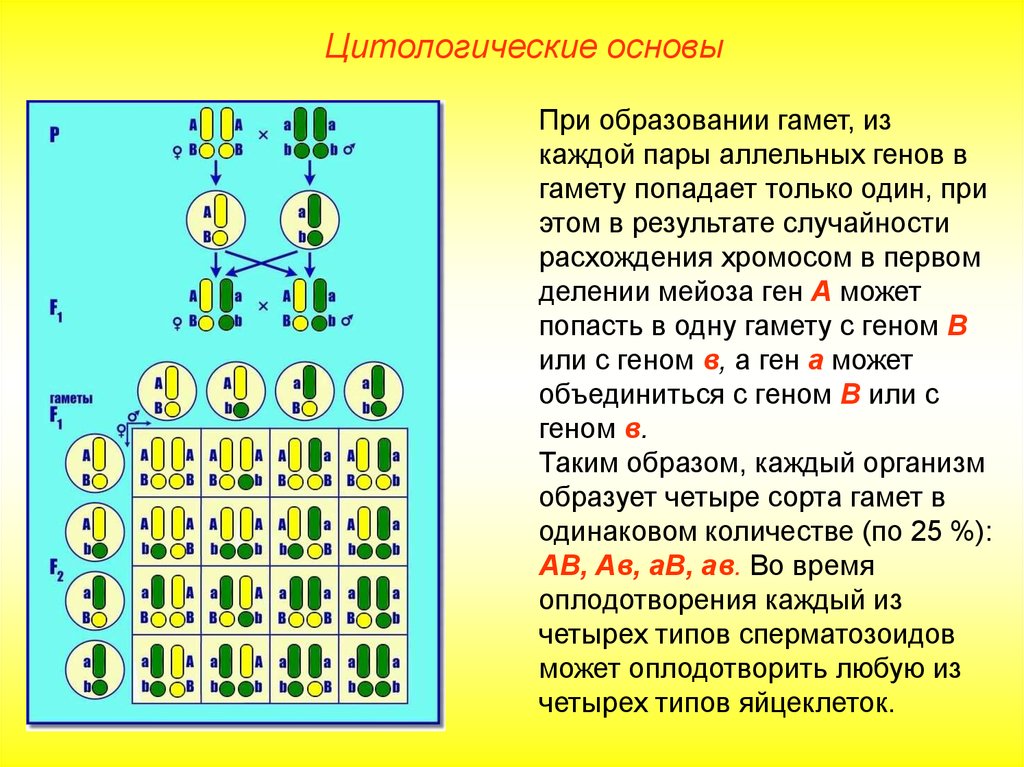
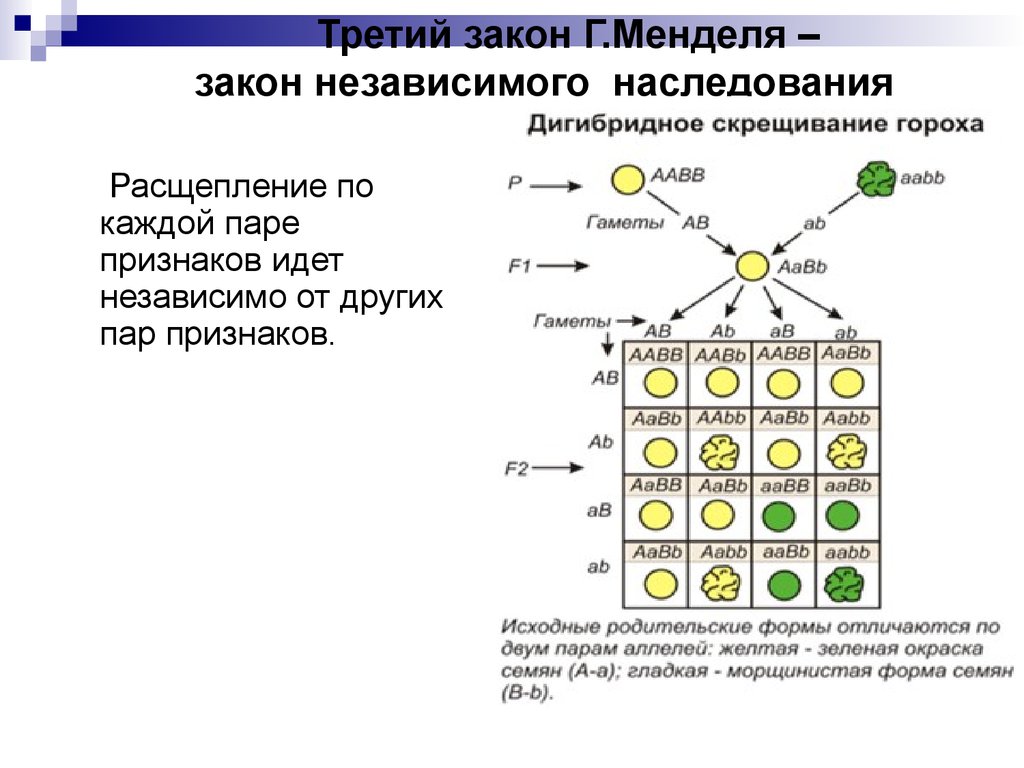
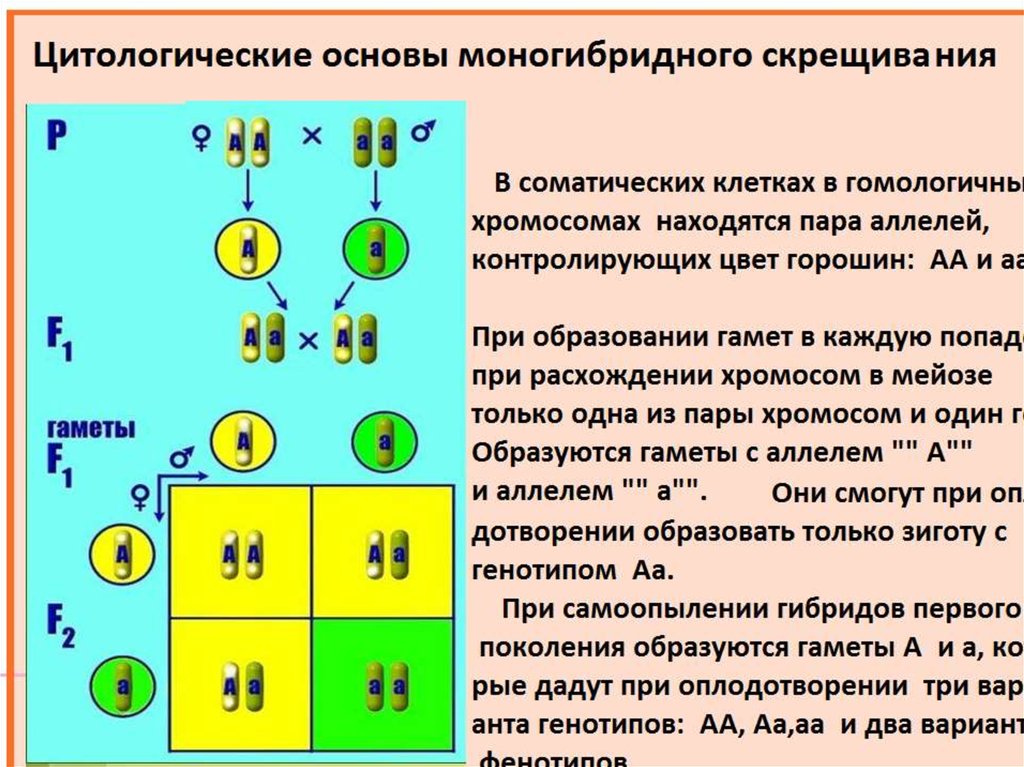
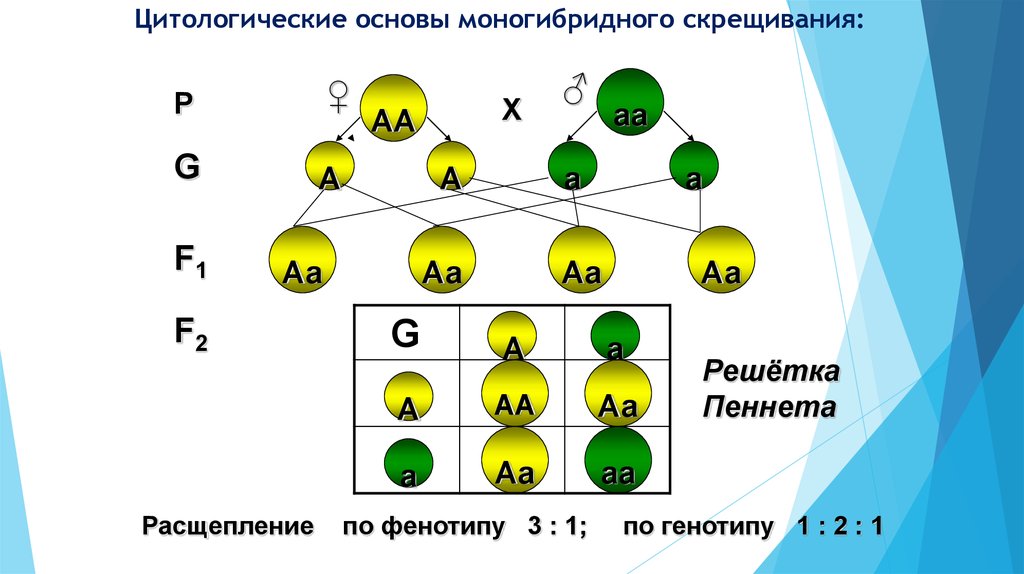
В линии с фиксированным геном (принимая во внимание данный локус) все особи имеют сходный генотип. В то же время, если в линии фиксированы все локусы, наступает генетическая сбалансированность и образуется инбредная линия
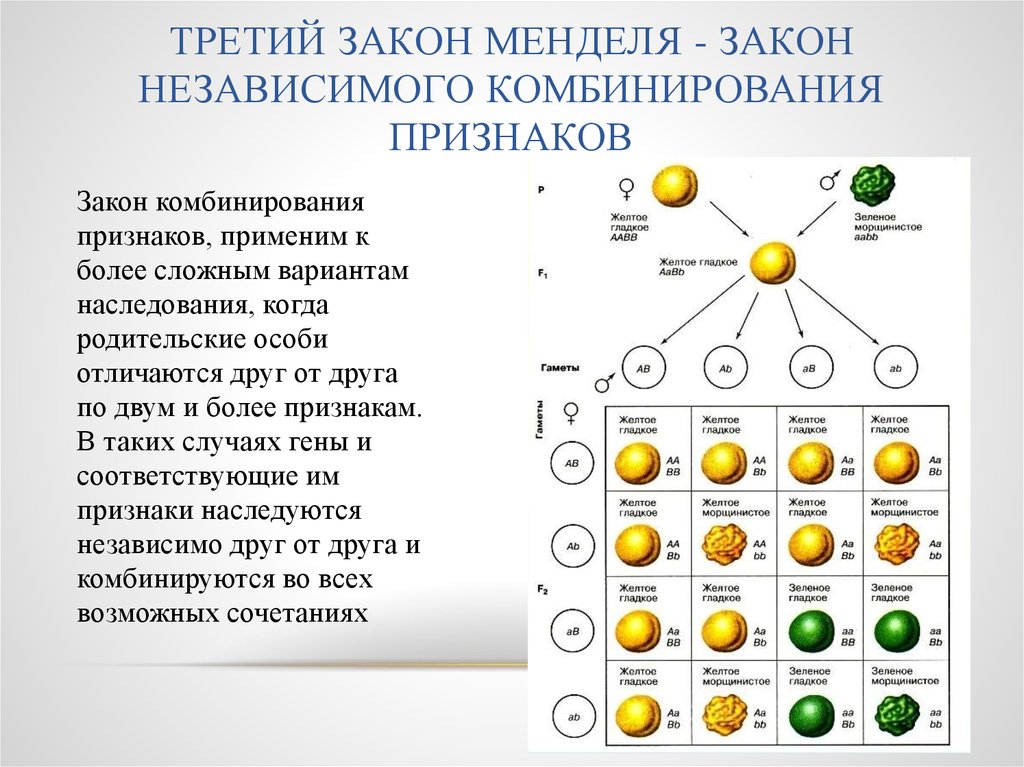
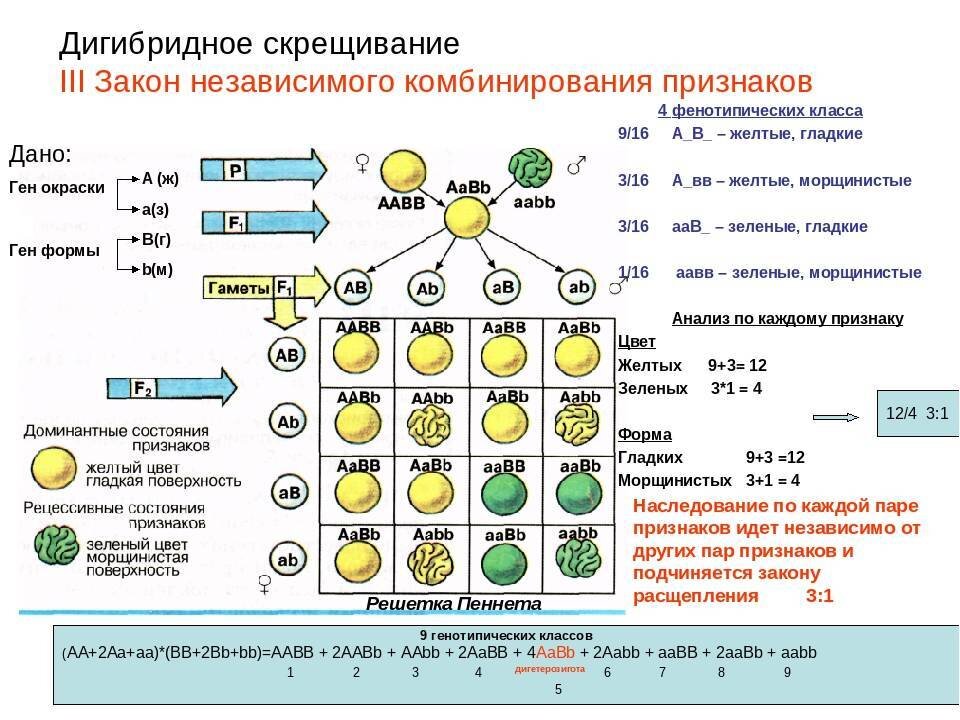
Фиксирование гена можно рассмотреть на конкретном примере. Если любая из 100 популяций начинает формироваться из двух гетерозигот (АаxАа), то потомство первого поколения будет состоять из трех генотипов: АА, Аа, аа в соотношении 1:2:1. В следующем поколении при наличии только двух родителей существует вероятность появления комбинаций из трех указанных генотипов, однако вероятность возникновения определенной комбинации зависит от частот проявления генотипов (табл. 5.2).
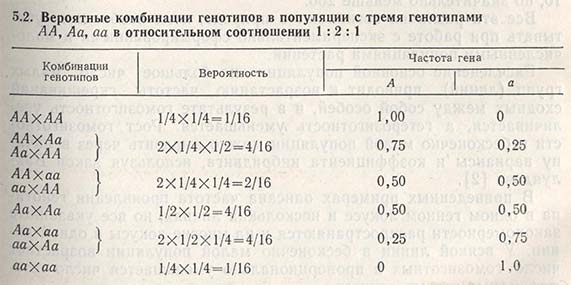
Вероятность случайного подбора обоих родителей с генотипом АА составляет 1/16, с генотипом аа — также 1/16. Это означает, что уже в первом поколении в 6 популяциях из 100 наверняка произойдет фиксация гена АА и в других 6 популяциях — фиксация гена аа. В остальных 88 популяциях, возможно, останутся следующие комбинации генотипов: ААxАа, ААxаа, AаxAа, Ааxаа. Три из этих комбинаций могут вновь дать популяции, гомозиготные по обоим аллелям, даже при условии, что в следующем поколении появится уже 15 таких популяций (15/100 = 15%). В последующих поколениях процесс будет продолжаться до тех пор, пока число популяций, достигших состояния фиксации, не станет равным 0,5N (где N — число родителей, использованных для получения поколений), т.е. при наличии двух родителей и 100 популяций 0,5N = 100. Это означает, что все 100 популяций после бесконечно большого числа поколений будут состоять из групп, достигших состояния фиксации, а частоты проявления их генов будут поделены между группой АА и группой аа пропорционально величинам р и q в основной популяции.
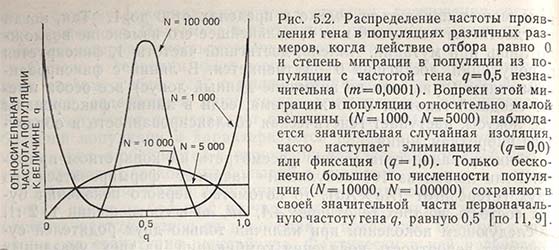
Таким образом, скорость фиксации гена зависит не только от частоты его проявления, но и от числа родителей в популяции. Если N равно 100 000, а первоначальная частота гена 0,5, то после продолжительного периода наступает случайная изоляция, хотя и весьма незначительная. При одинаковой частоте проявления гена и малой величине популяции от N = 1000 происходит быстрая фиксация генов и элиминация аллеля (рис. 5.2).
Число родителей имеет большое значение для установления генетической изоляции и выражается как эффективная величина популяции (Ne). Этот показатель отличается от действительной величины, поскольку не все члены популяции являются родителями и дают потомство. Если из 1000 особей только у 400 пар происходит скрещивание, эффективная величина популяции равна 800. При соотношении полов, не равном 1:1, наступает уменьшение эффективной величины популяции: когда на 200 особей женского пола приходится 10 особей мужского пола, Ne превышает 10, но значительно меньше 200. Все эти различия возникают в природе, и их необходимо учитывать при работе с экспериментально сформированными и малочисленными популяциями растений.
Расчленение основной популяции на большое число малых групп (линий) приводит к возрастанию частоты скрещиваний сходных между собой особей, и в результате гомозиготность увеличивается, а гетерозиготность уменьшается. Рост гомозиготности в бесконечно малой популяции можно выразить через величину вариансы и коэффициента инбридинга, используя закон Вахлундова.
В приведенных примерах описана частота проявления генотипа в одном генном локусе и нескольких линиях, но все указанные закономерности распространяются и на многие локусы в одной линии. У всякой линии в бесконечно малой популяции возрастает число гомозиготных и пропорционально уменьшается число гетерозиготных генных локусов.
Рассмотренный в главе общий характер дисперсных процессов показывает, что дифференциация популяций на уровне подлиний вследствие случайной изоляции и фиксации генов приводит к сокращению масштабов изменчивости, росту гомозиготности и сбалансированности подлиний. Полной гомозиготности бесконечно малая популяция растения-перекрестника достигает в процессе инбридинга, если он возможен у данного вида.
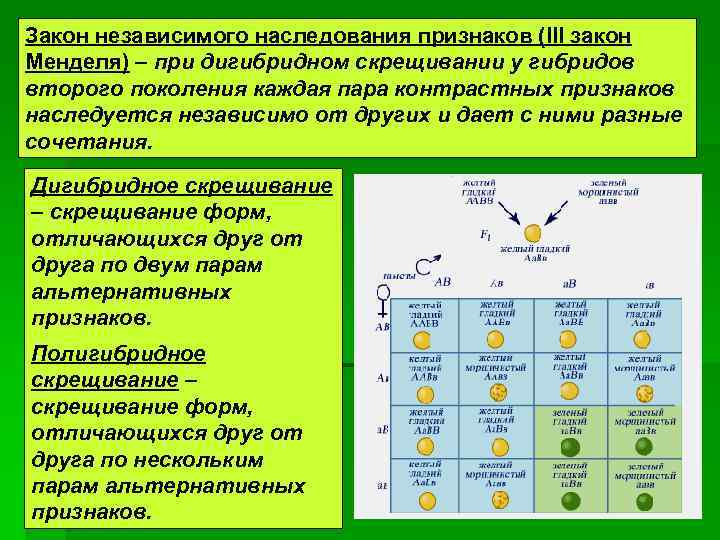
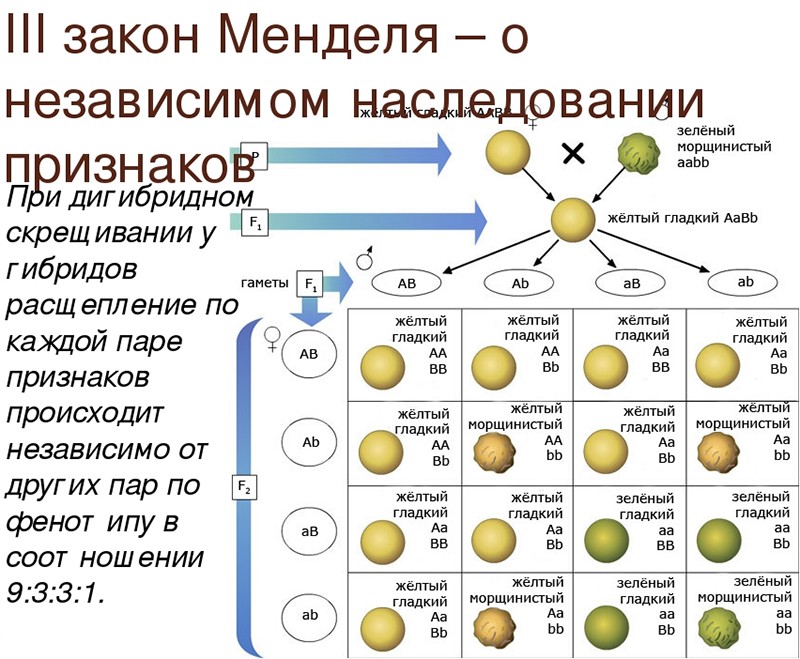
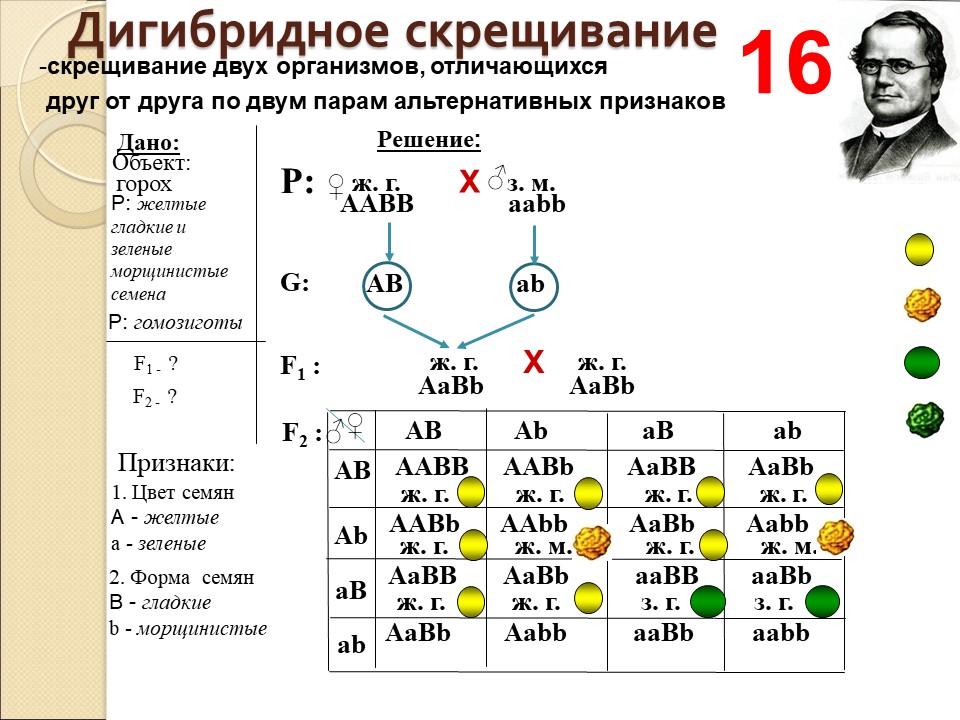
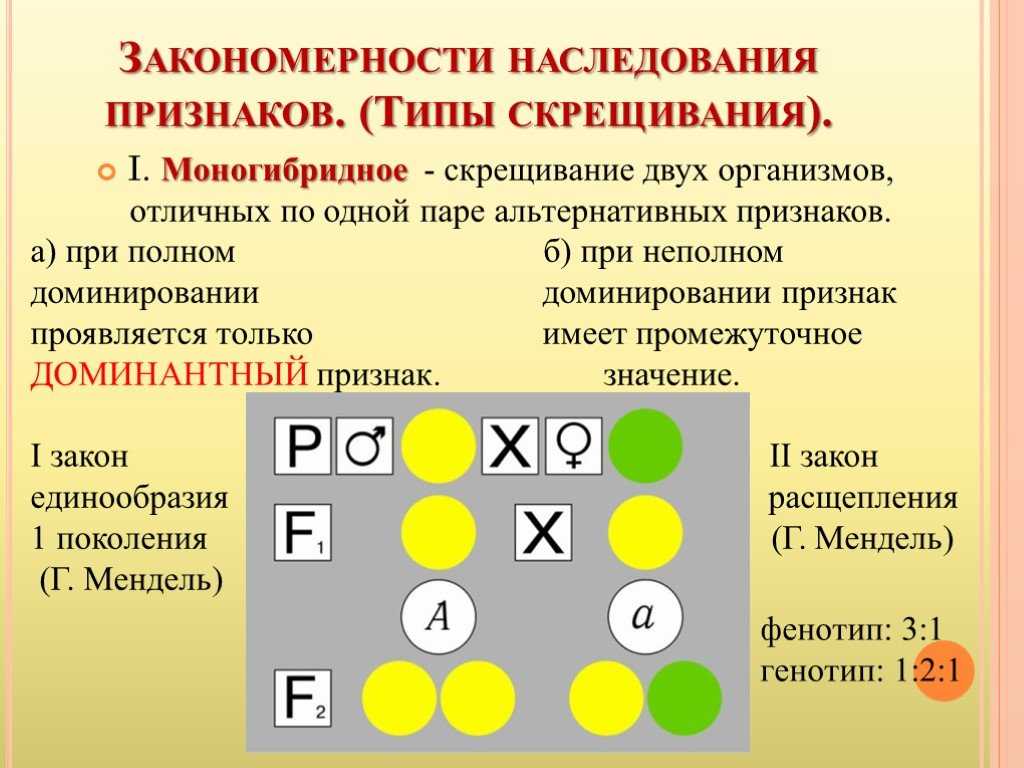
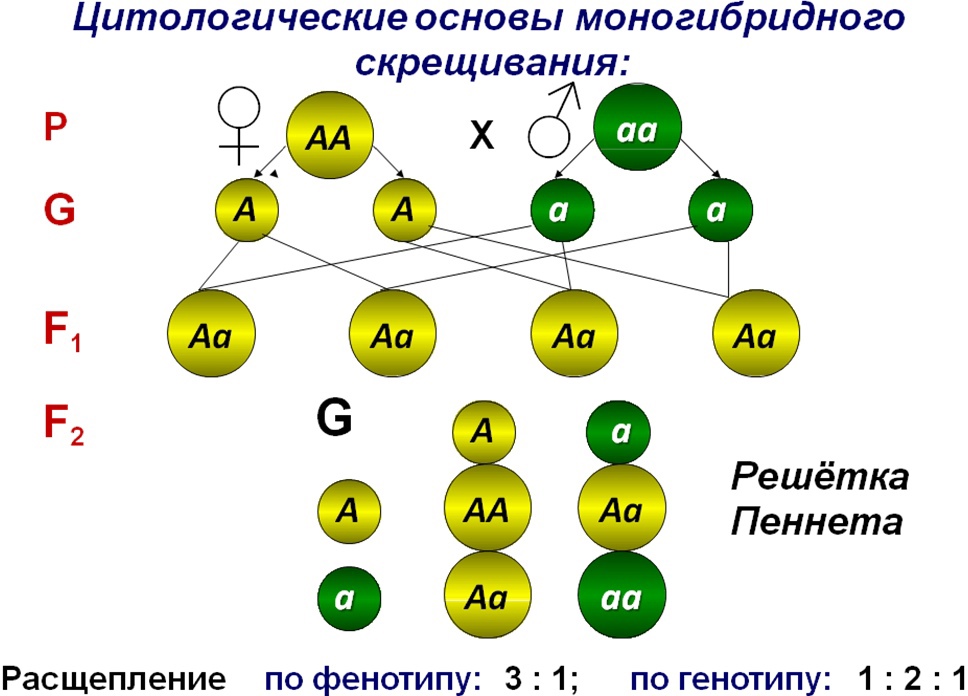
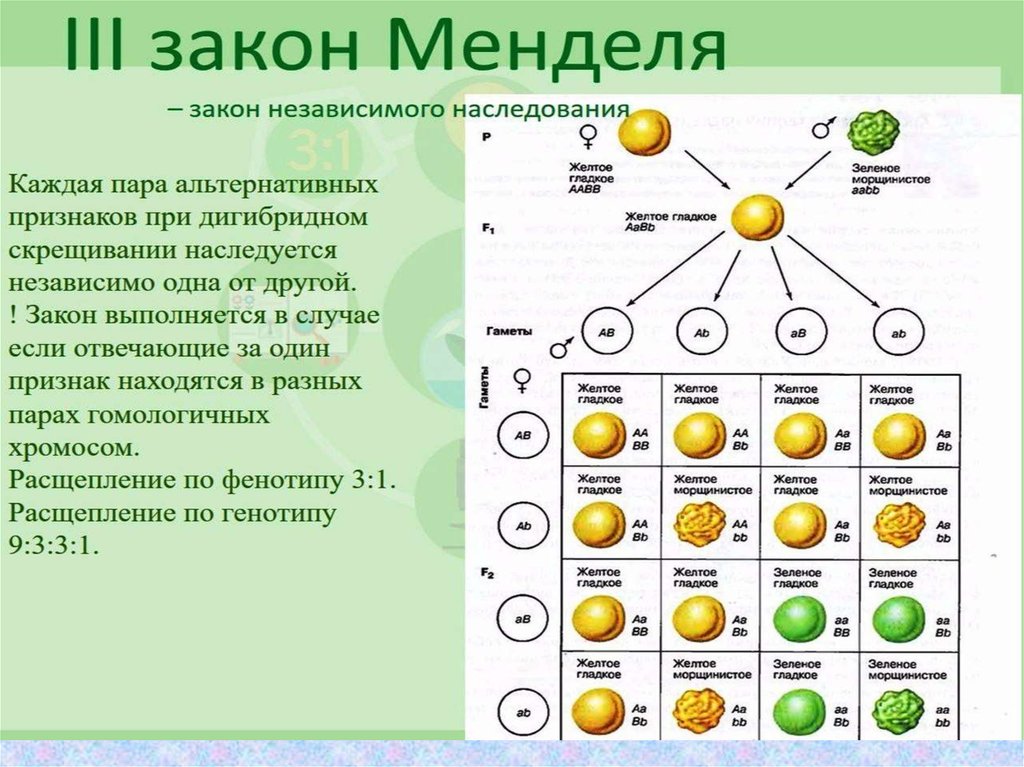
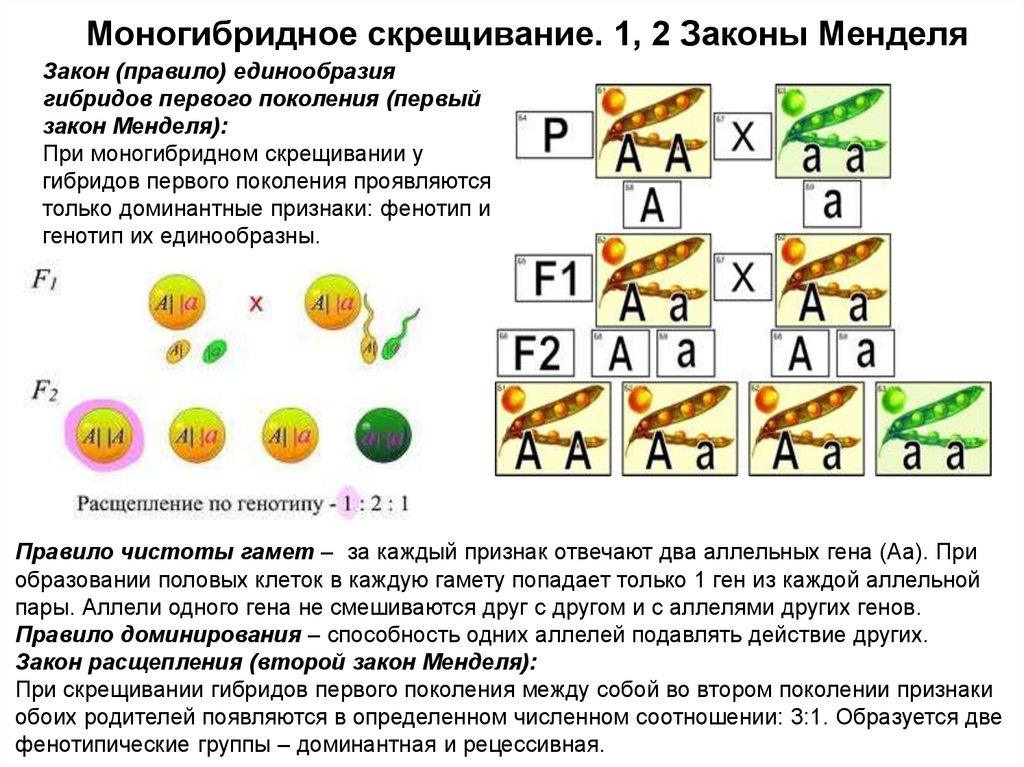
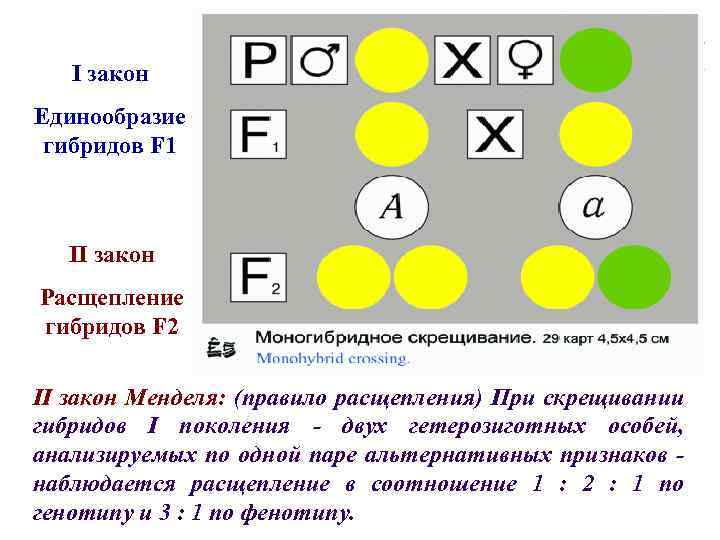
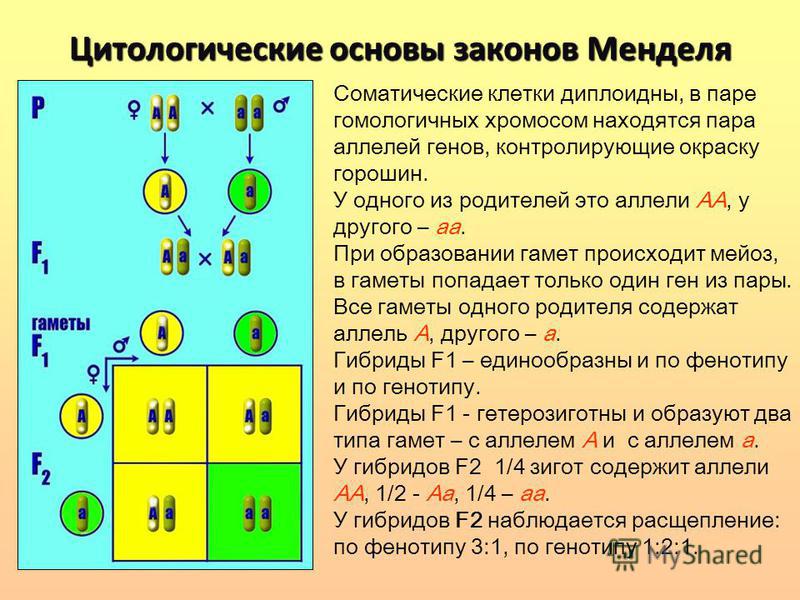
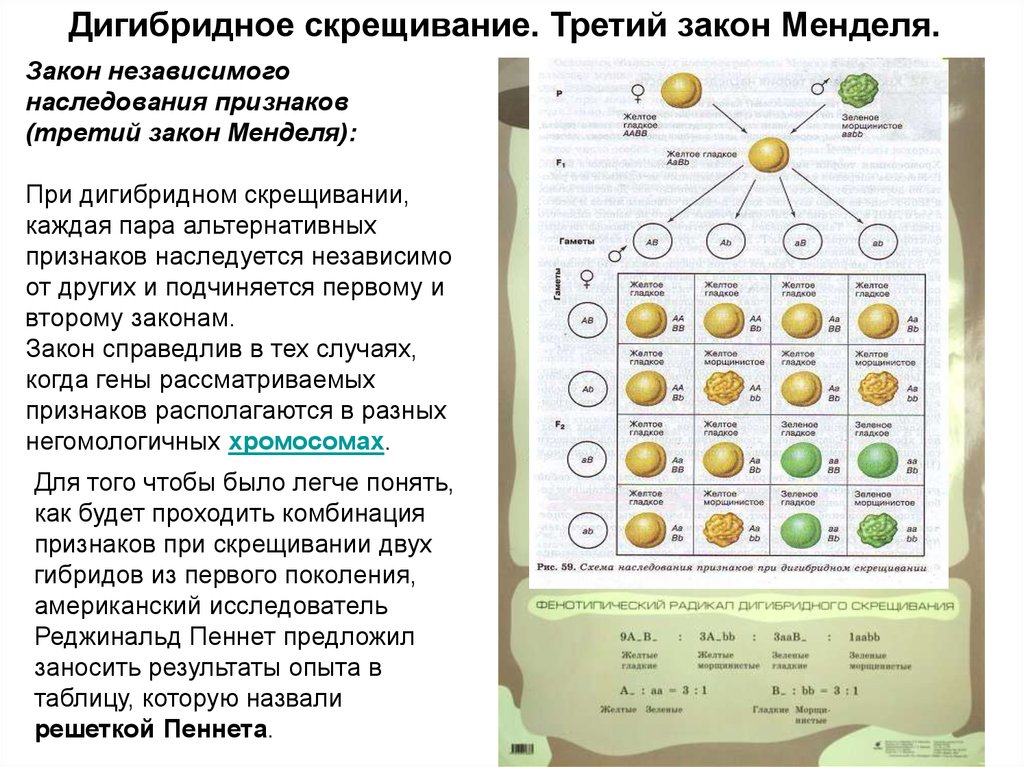
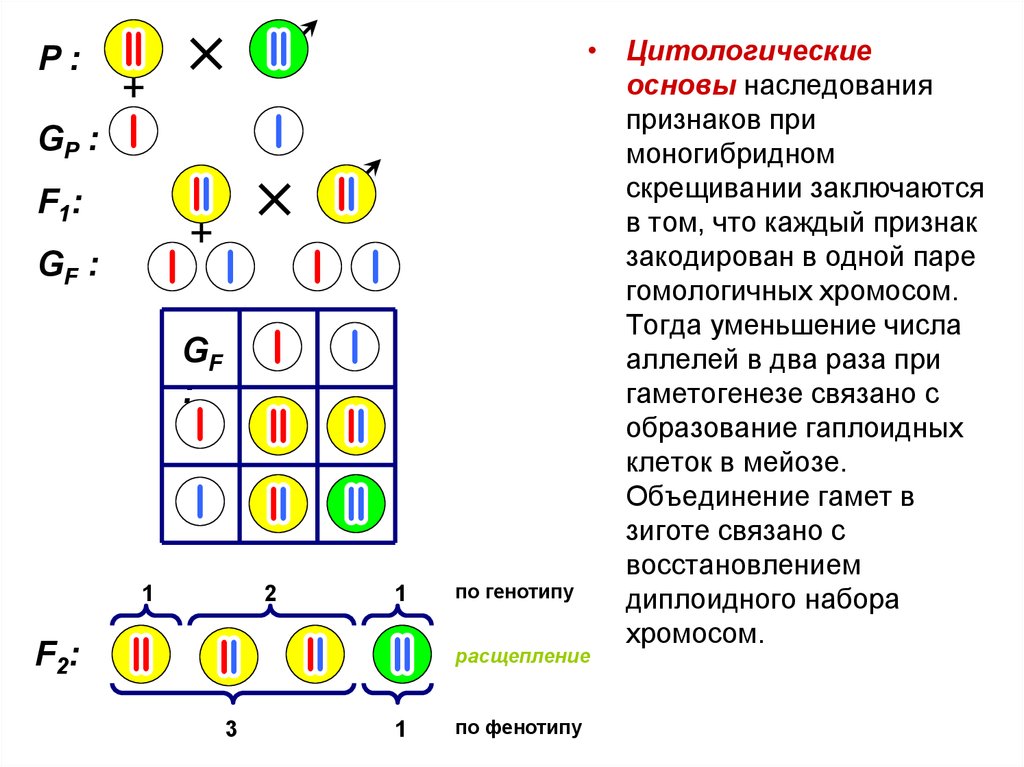
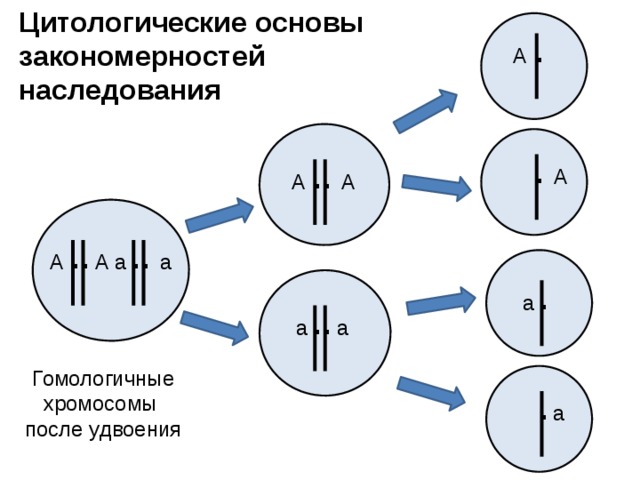
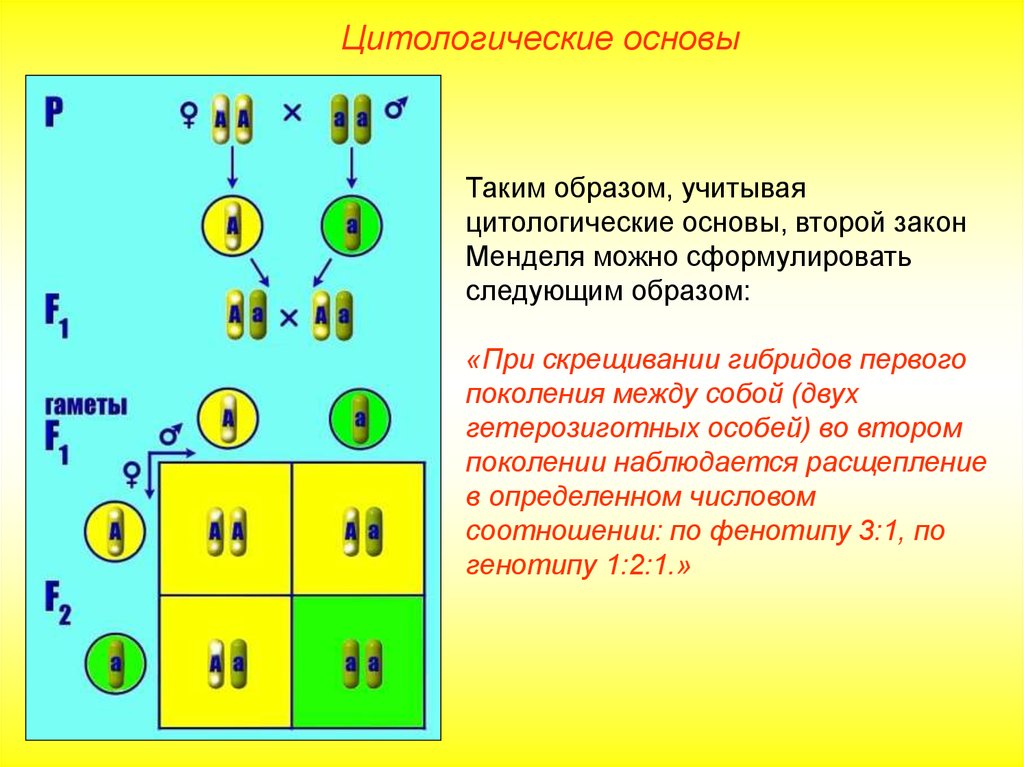
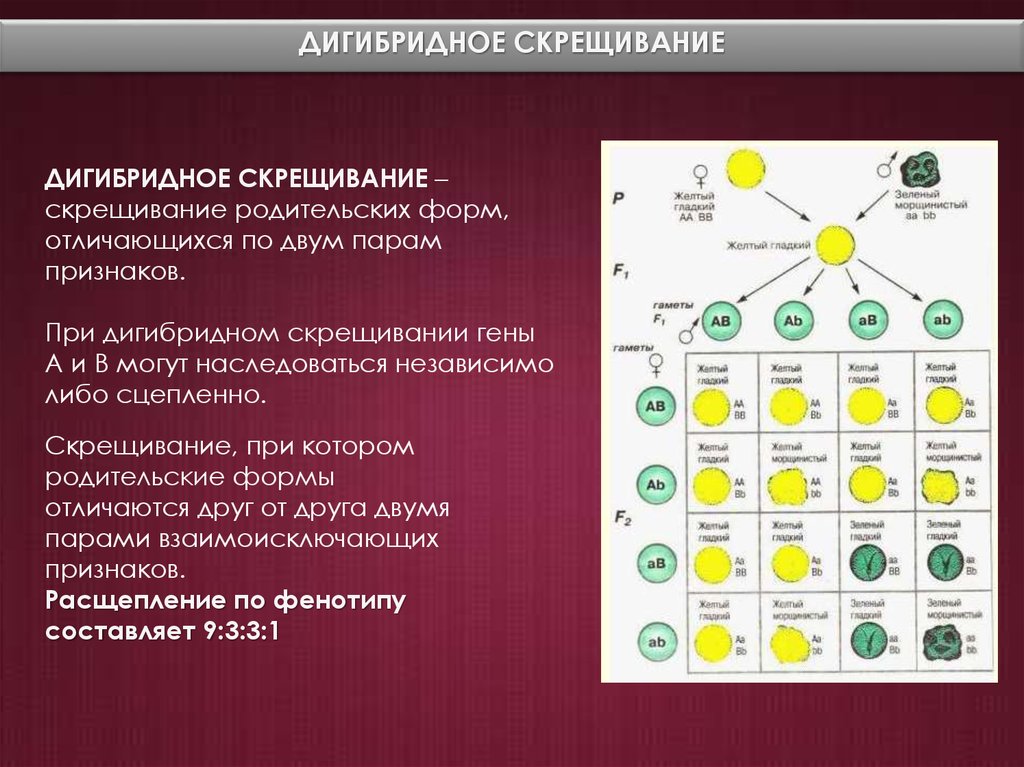
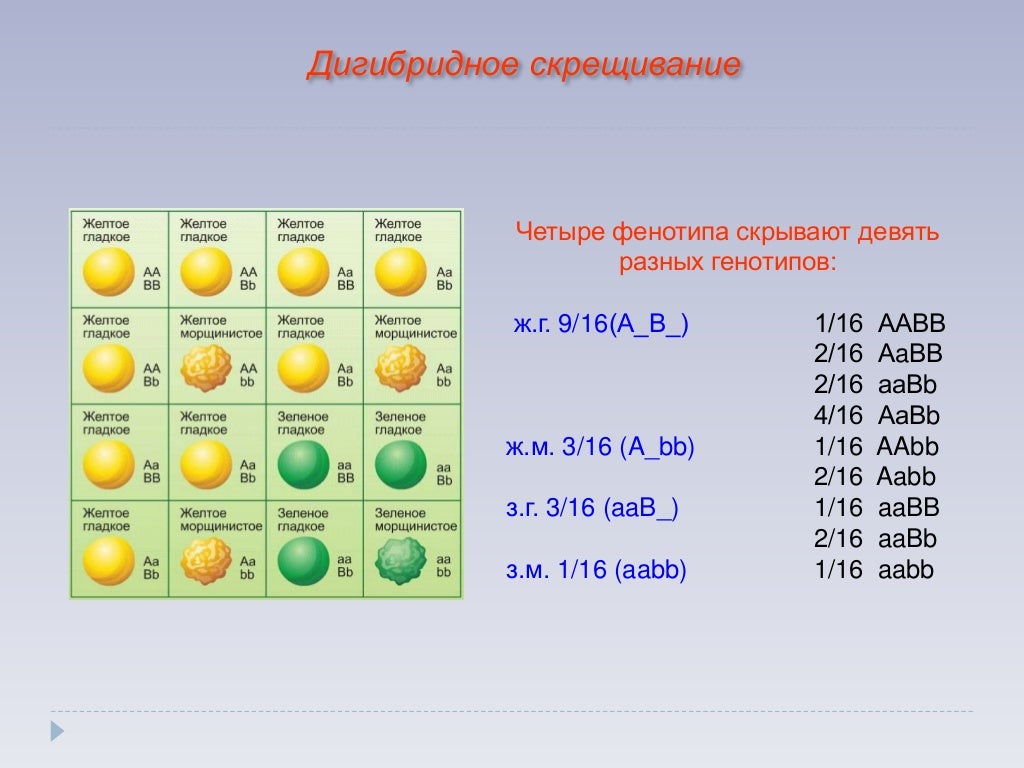
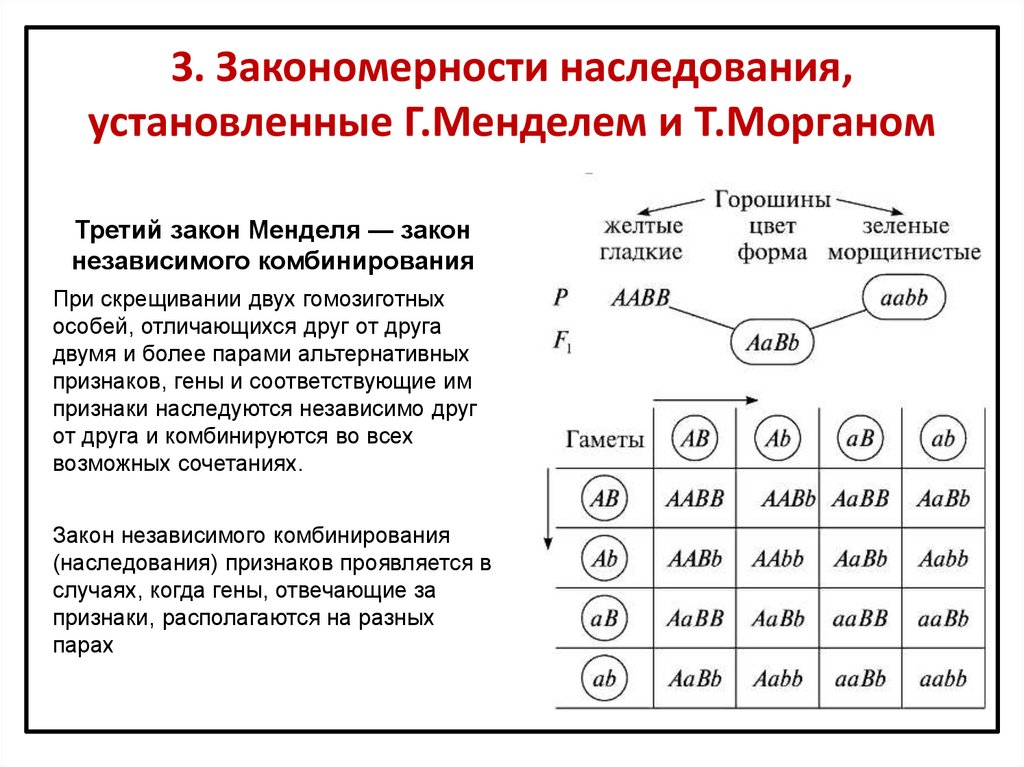
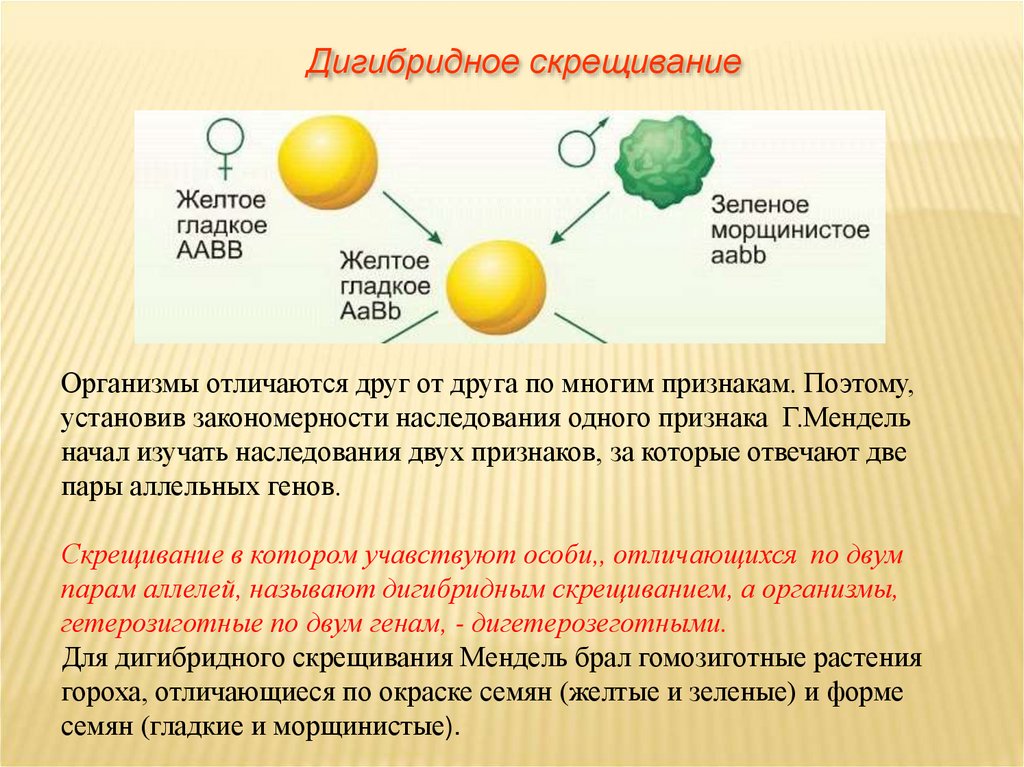
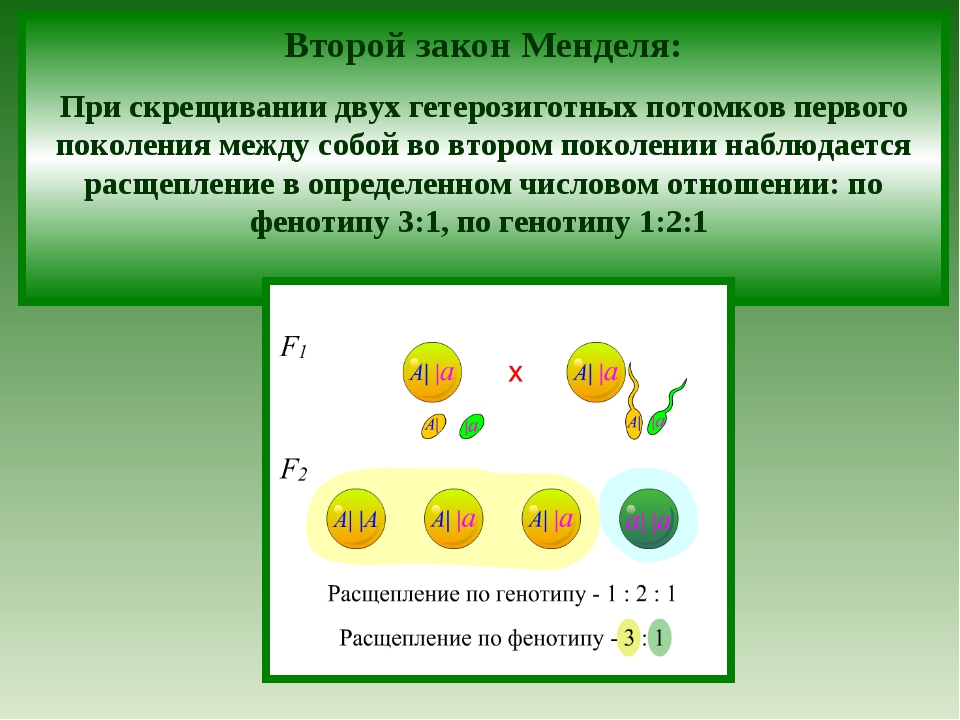
Второй закон Менделя — в чем суть
Рассматриваемые принципы легли в основу классической генетической науки и биологии. После представления описанные закономерности получили объяснение в форме результата важных молекулярных механизмов наследственности.
В источниках, опубликованных на русском языке, упоминается три таких закона. Однако открытие «первого закона» не принадлежит Менделю. Большую ценность для развития генетики представляет «гипотеза чистоты гамет».
Рассмотрим второй закон. В рамках эксперимента Г. Мендель наблюдал за самостоятельным опылением гибридов из первого поколения. В результате сформировались гибриды во втором поколении. Эти организмы приобрели, кроме доминантного, рецессивный признак. Краткие итоги подобного опыта описаны в таблице ниже:
В результате исследования табличных данных можно сформулировать ряд очевидных выводов:
- Отсутствует единообразие гибридов во втором поколении. При этом гибриды частично обладают одним (доминантным) признаком, частично имеют другой (рецессивный) признак из альтернативной пары.
- Численность гибридов, которые несут доминантный признак, примерно в три раза превосходит количество гибридов, несущих рецессивный признак.
- Исключается исчезновение рецессивного признака у гибридов первого поколения. Можно наблюдать подавление данного признака и его проявление у гибридов второго поколения.
При наблюдении расщепления у гибридов можно заключить, что явление имеет не случайный характер, а определено численными закономерностями. Основываясь на данном факте, Мендель пришел к следующему выводу: в результате скрещивания гибридов первого поколения в потомстве протекает расщепление признаков согласно конкретному количественному соотношению.
Скрещивание растений
Плоды земляники садовой. Предположительно возникла в результате скрещивания земляники чилийской и земляники виргинской
В скрещивании растений в науке опытным путем достигнуты различные варианты гибридизации.
Анализирующее скрещивание — скрещивание гибридной oсоби с особью, гомозиготной по рецессивным аллелям, то есть «анализатором».
Беккросс — скрещивание гибридов F1 с одной или обеими родительскими формами.
Внутривидовое скрещивание — скрещивание организмов, относящихся к одному и тому же виду.
Диаллельное скрещивание — cкрещивание, при котором испытываемые линии или сорта скрещиваются во всех возможных комбинациях.
Инконгруэнтное скрещивание (трудноудающееся) — это отдалённые скрещивания между организмами, имеющими разные наборы хромосом или разное их число. Могут быть межвидовыми или межродовыми.
Конгруэнтное скрещивание — это скрещивание организмов с совместимыми наборами генетических хромосом.
Наcыщающее cкрещивание — это многократное возвратное скрещивание гибридов или форм с одной из исходных родительских форм.
Ступенчатое скрещивание — это скрещивание, при котором последовательно скрещивают несколько разных родительских форм .
Топкроссы — это cкрещивания, когда ряд исходных сортов скрещивают с определённым набором других сортов.
Простые скрещивания — это скрещивания, при которых разные родительские формы участвуют только в одной комбинации.
Реципрокные скрещивания (взаимные скрещивания) — это cкрещивания между двумя формами, когда каждая из них в одном случае используется в качестве материнской, а в другом — в качестве отцовской.
Преимущества скрещивания
Процесс гибридизации — скрещивания растений — направлен на получение сортов растений, обладающих выигрышными свойствами родительских сортов, таких как:
- Высокая урожайность
- Устойчивость к заболеваниям
- Устойчивость к морозам
- Устойчивость к засухе
- Короткие сроки созревания
- Лучшее плодоношение
К примеру, если у отцовского и материнского сорта растений проявляется устойчивость к разным заболеваниям, то полученный гибрид возможно унаследует стойкость к обеим болезням. Гибридные сорта растений обладают лучшей жизнестойкостью, они меньше подвержены повреждениям от перепадов температуры, влажности, изменений климатических условий, чем их негибридные собратья.


Бесконечно малая популяция
Рассмотренные явления и закономерности касаются бесконечно большой по численности популяции (из нескольких тысяч особей), в которой частота проявления генов и генотипов остается неизменной из поколения в поколение, если на нее не воздействуют извне отбор, мутации и миграции. Эти воздействия имеют систематический и постоянный характер независимо от их направленности в сторону фиксации генов или в сторону достижения состояния равновесия. Поэтому в естественных популяциях можно предусмотреть направленность указанных воздействий.
Однако в селекции чаще всего приходится работать с небольшим числом особей, т.е. с малой (первичной, или исходной) популяцией, содержащей лишь часть генетической изменчивости. Такая популяция представляет собой лишь модель бесконечно большой популяции и характеризуется рядом отступлений и специфических особенностей в отношении тех закономерностей, которые управляют бесконечно большой популяцией.
В бесконечно малой популяции отбор уклоняющихся генотипов (точнее гамет из большой по численности популяции) ведет к изменению частоты генов из поколения в поколение, поэтому популяция эта структурно неустойчива. Фактически в ней разворачиваются дисперсные процессы, приводящие к: 1 — разделению популяции на подпопуляции, группы, линии; 2 — уменьшению генетической изменчивости и возрастанию уровня гомозиготности.
В подпопуляциях или группах все активнее идет скрещивание сходных между собой особей, т.е. инбридинг, поэтому внутри групп наблюдается уменьшение генетической изменчивости, фиксация одних и элиминация других генов и увеличение гомозиготности. Таким образом, наступает разделение между группами или линиями, которое Райт назвал случайным дрейфом (random drift).
Образование различных местных популяций у растений-самоопылителей — пример действия дисперсных процессов, происходящих в естественных популяциях. Так, по данным Бригера, в Южной Америке встречаются многочисленные местные популяции кукурузы, которые в сильной степени отличаются друг от друга, хотя при этом в пределах отдельной популяции преобладает значительная выравненность. Подобная картина характерна и для югославских местных форм ржи, особенно в горных районах: под влиянием элементов рельефа, разделяющих поля, местные популяции разбились на ряд малых популяций, внутри которых все чаще и чаще происходит скрещивание сходных между собой особей. Это привело к повышению уровня гомозиготности в популяциях и одновременно к морфологической их выравненности. Так, местная рожь из Лике фенотипически однородна, но в значительной степени отличается от местной ржи из Гламоча и в еще большей степени от местной ржи из Невесиньи.
Подобные или сходные дисперсные процессы протекают и у других видов-перекрестников: злаковых трав, бобовых и т.д. Поэтому местные популяции представляют немалый интерес для дальнейшей работы по селекции растений.
Несовместимость
Для обеспечения перекрестного опыления, особенно у обоеполых цветков, в ходе эволюции возникли различные барьеры, препятствующие самоопылению. Существует несколько причин автостерильности или несовместимости, к которым относятся:
протандрия — когда в одном и том же цветке тычинки с пыльниками созревают раньше пестиков и пыльца попадает не на рыльца данного пестика, а на различные другие цветки;
протогиния — когда пестик созревает раньше тычинок с пыльниками, что также не может привести к оплодотворению в одном и том же цветке;
геркогамия — разделение на мужские и женские генеративные органы, при котором на одном и том же растении не может произойти самооплодотворение.
Наряду с этими морфологическими и механическими механизмами, обусловливающими перекрестное оплодотворение, причины несовместимости у черешни, огромного числа сортов яблони, груши и миндаля, сливы, вишни и маслины и далее у ржи, сахарной свеклы, люцерны, некоторых видов кормовых злаковых трав, табака и т.д. имеют генетическую и физиологическую природу. Льюис разделяет несовместимость у растений на две группы — гетероморфную и гомоморфную.
Гетероморфная несовместимость
Система несовместимости, основанная на различии в длине тычинок и пестика, называется гетеростилией. Тип цветка с длинным пестиком и пыльниками на коротких тычиночных нитях Льюис назвал pin (гвоздь, игла); тип цветка с коротким пестиком и длинными тычиночными нитями был назван thrum (сережка, букет).
Генетическая конституция типа pin — ss, типа thrum — Ss, при этом 5 полностью доминирует над s (рис. 5.1).
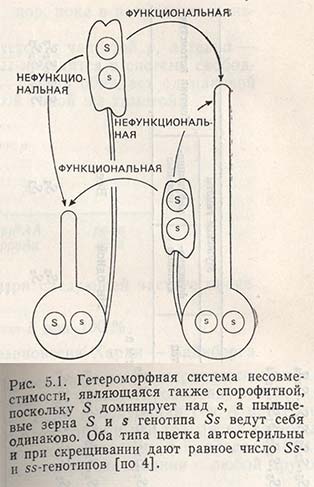
Оплодотворение происходит только между следующими генотипами:
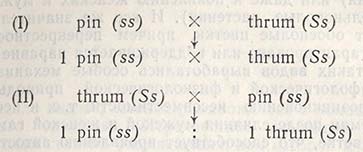
Комбинации pin X pin и thrum X thrum несовместимы, т.е. генотипы типа SS не возникают (рис. 5.1).
Гомоморфная несовместимость
Гомоморфная несовместимость разделяется на гаметофитную и спорофитную.
Гаметофитная несовместимость была впервые установлена у Nicotiana sanderae Истом и Мангельсдорфом. Позднее ее обнаружили у клевера красного и белого и некоторых других кормовых бобовых. Функциональность пыльцевых зерен зависит от ряда аллелей в одном локусе, обозначенных как s1, s2, s3, … sn. Если в клетках присутствует тот же ген, что и в пыльцевом зерне, то пыльцевая трубка не может развиваться в пестике и оплодотворения не происходит. Оно имеет место только, когда пыльца несет гены, отличные от находящихся в пестике и яйцеклетке. Способность пыльцевого зерна к оплодотворению зависит от его собственного генотипа. Следовательно, при скрещивании растений с различной генетической конституцией может развиваться: полная несовместимость, совместимость половины всех пыльцевых зерен и полная совместимость (табл. 5.1).
Спорофитная несовместимость. Эта система несовместимости также обусловлена рядом аллелей в одном локусе, по в отличие от гаметофитной системы в данном случае функциональность пыльцевых зерен определяется генетической конституцией растения, которое их продуцировало (см. табл. 5.1).
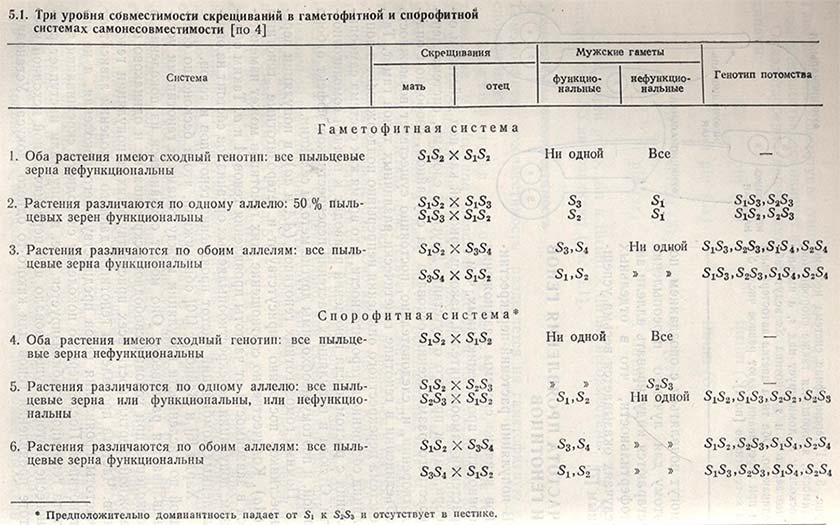
Несовместимость создает сложности, поскольку у ряда видов невозможно получить инбредные линии для использования их на гетерозисной основе. И все же несовместимость стремятся применять главным образом в селекции овощных и цветочных культур. Об этом пойдет речь в главе, посвященной методам отбора.
Особенно надлежит учитывать явление несовместимости при возделывании овощных культур. Оно может обусловливать слабое оплодотворение и приводить к снижению урожаев, следовательно, нужно обязательно добиваться хорошего переопыления сортов. При неблагоприятных погодных условиях насекомые-опылители могут появляться с опозданием, поэтому для лучшего переопыления стараются индуцировать аллели автофертильности, что в отдельных случаях оказывается весьма успешным.
Скрещивание животных
Родительские формы яичного кросса «Хай-Лайн коричневый» Щучинской птицефабрики на выставке «Белагро-2019»
В отличие от чиcтопородного разведения при скрещивании спаривают животных различных пород. Животных, полученных от скрещивания, называют помеcями.
Скрещивание применяется в животноводстве с древнейших времён. В итоге скрещивания повышается гетерозиготность получаемых животных, что часто сопровождается появлением такого биологического явления, как гетерозиc (значительное превосходство помеcей над лучшей из исходных пород), который в пользовательном животноводстве играет большую роль, иногда и решающую.
Скрещивание — это самый эффективный метод быстрого изменения наследственных признаков животных и создания новых высокопродуктивных пород. Успех скрещивания зависит от многих факторов: умелого выбора исходных пород, штаммов, сортов; цели и вида скрещивания; подбора лучших производителей, проверенных по качеству потомства; условий кормления и содержания полученного помесного поголовья.
В зависимости от поставленной цели выделяют основные виды скрещивания: воспроизводительное (скрещивание, направленное на выведение новой породы); поглотительное (для преобразования худших пород в лучшие); промышленное (для использования гетерозиса помесей первого поколения); переменное (для удержания гетерозиса в ряде поколений); вводное (частичное улучшение одной породы путем однократного скрещивания с животными другой породы).
Примечания
- . Дата обращения: 21 августа 2023.
- . Большая советская энциклопедия. Дата обращения: 28 августа 2023.
- ↑ Батанов С. Д., Старостина О. С., Атнабаева Н. А. Промышленное скрещивание как основной резерв получения высококачественной говядины в молочном скотоводстве // Молочное и мясное скотоводство. — 2022. — № 5. — С. 13—16.
- Левахин В.И. Эффективность промышленного скрещивания в скотоводстве // Молочное и мясное скотоводство. — 2002. — № 1. — С. 9—11.
- Стрельцов В. А. Организация племенной работы со свиньями на промышленных фермах и комплексах не имеющих собственных племферм // Вестник Брянской государственной сельскохозяйственной академии. — 2011. — № 5. — С. 72—76.
- . Мegaogorod.com. Дата обращения: 21 августа 2023.
Что такое эволюция — понятие
Определение
Эволюция является биологическим процессом, соотносящимся с временными изменениями, в ходе которых набор генов видоизменяется на уровне ДНК, в результате чего образуется множество потомков.
Понятия эволюции и естественного отбора часто пересекаются.
Определение
Процессом эволюционирования называют историческое формирование мутаций.
Осторожно! Если преподаватель обнаружит плагиат в работе, не избежать крупных проблем (вплоть до отчисления). Если нет возможности написать самому, закажите тут
Естественный отбор — наиболее важный из механизмов, за счет которого возникают мутации. В современном мире область эволюционной биологии получила активное развитие, было изучено большое количество данных по теме эволюции.



Ученым удалось исследовать исторические эволюционные изменения, которые были выведены непосредственно при анализе окаменелостей, изучения филогении и процесса эволюционных изменений, в том числе, эффекта естественного отбора. Известно, что сильный отбор ускоряет эволюцию сильнее, чем было предположено Дарвином.
Результатом современных исследований является определение возможности для проведения эволюционных опытов в реальном времени. Последние события во многих научных областях, таких как молекулярная биология и биология развития, существенно увеличили спектр имеющихся знаний и послужили доказательством того, что эволюция занимает центральное место в понимании биологического разнообразия.
Кратко, эволюция представляет собой процесс последовательного естественного развития. Для него характерны следующие черты:
- природа и социальные системы изменяются необратимо и постепенно;
- живые организмы адаптированы к окружающей среде и внешним факторам, что выражается в борьбе за существование;
- активная оболочка планеты в виде биосферы преобразуется;
- старые генетические популяции отмирают, возникают новые, а также наблюдаются мутации в животном и растительном мире.
Примечание
Важной особенностью эволюционного процесса является то, что он протекает естественно, планомерно, исключаются бурные всплески и скачки. Данные факты отличают его от революционных преобразований
Основателями теории биологической эволюции являются Мендель, Ламарк, Дарвин. Вместе с тем, современная философская наука все чаще оперирует понятием социальной эволюции, которая характеризует изменения в общественной жизни.