
Система компактизации хромосом
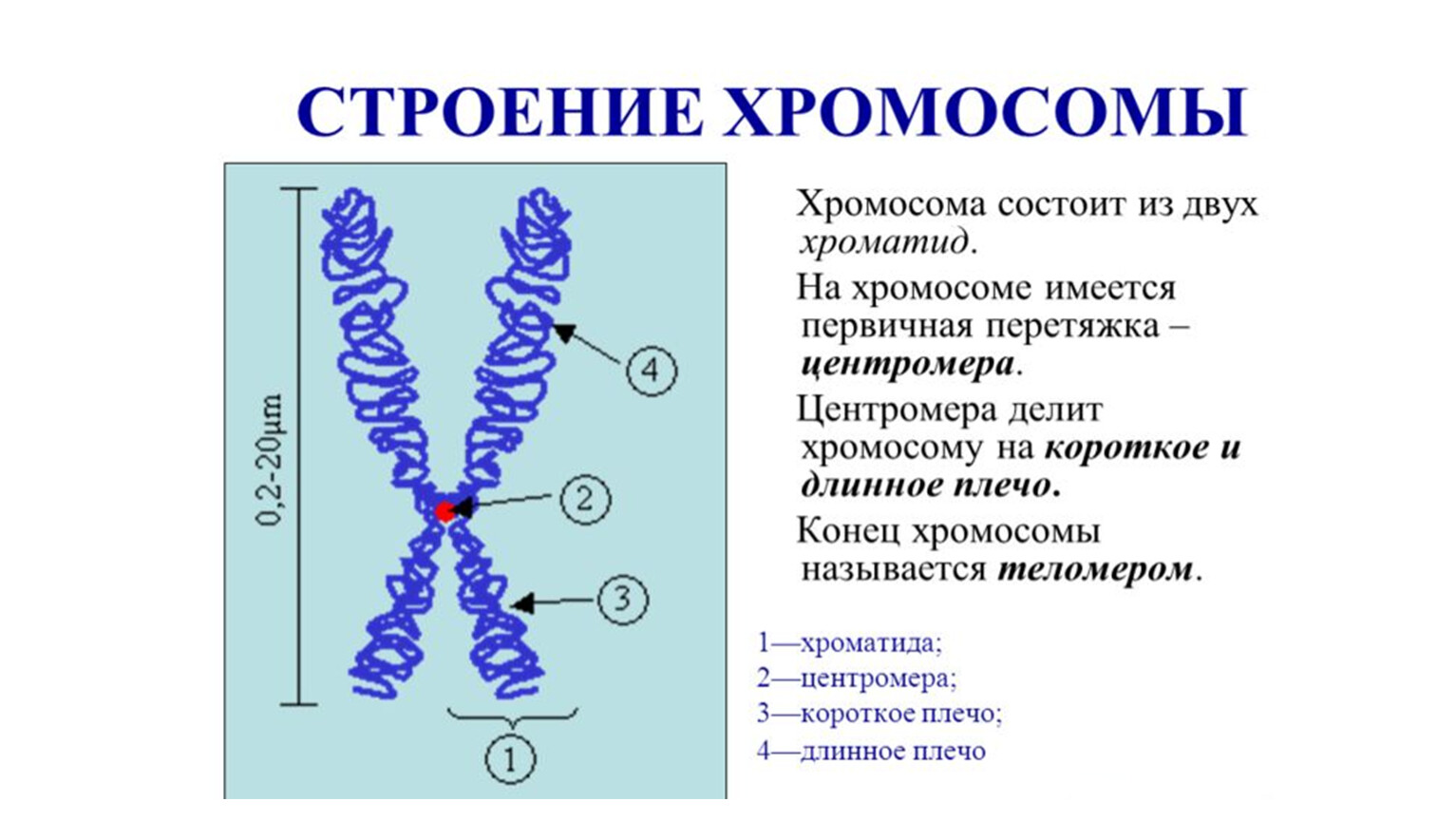
Хромосомы состоят из двух плачей, которые соединяются хроматидой. Суммарная длина ДНК внутри клеток человека составляет приблизительно 2 метра. Диаметр ядра клетки составляет приблизительно 7 мкм. При учете того факта, что каждая хромосома представлена отдельной молекулой ДНК, то компактизация ДНК составляет более шести тысяч раз. Подобная укладка происходит в несколько уровней:
- Первый уровень компактизации хромосом называется нуклеосомным. Если хроматин находится под действием нуклеазы, то ДНК распадается и состоит из регулярно повторяющихся структур. Этими структурами являются 8 молекул белков – гистонов, намотанных на октамер ДНК. Этот комплекс называется нуклеосомой. В ней гистоны образуют белковую основу-сердцевину, по поверхности которой располагается почти 2 витка ДНК. При этом между нуклеосомами остается линкер, участок, соединяющий две соседние нуклеосомы. Такой уровень компактизации ДНК часто называют «бусины на нитке». Буснки – это нуклеосомы и нитка является ДНК. Диаметр нуклеосомы около 11 нм. Такой уровень компактизации ДНК играет структурную роль и обеспечивает плотность упаковки ДНК;
- Второй уровень компактизации ДНК составляет 30 нм и имеет вид фибриллы. В митотических хромосомах можно обнаружить фибриллы, которые обладают диаметром 25 – 30 нм. При этом также можно наблюдать соленоидный тип нуклеосом в виде нити с витками и спиральным шагом около 10 нм. На каждом витке формируется фибрилла с центральной полостью. Хроматин в составе ядер таких клеток будет иметь фибриллы по 25 нм. Этот тип укладки ДНК обеспечивает ее сорокакратное уплотнение.
- Третий уровень компактизации ДНК представлен петлевыми доменами и называется хромомерным. Он считается уровнем высшего ранга. Внутри таких уровней специфические белки связываются с ДНК, образуя так называемые большие петли. В некоторых местах есть сгустки конденсированного хроматина, розетковидные образования, состоящие из многих петель 30 нм-фибрилл, соединяющихся в плотном центре.
Средний размер таких розеток достигает 100 – 150 нм. Розетки фибрилл хроматина называются хромомерами. Каждый такой хромомер состоит из нескольких нуклеосом или петель, которые связываются в одном центре. Хромомеры связываются друг с другом и некоторыми участками нуклеосомного хроматина. Это обеспечивает так называемую структурную компактизацию хроматина и организует функциональные хромосомные единицы, а именно репликоны и гены, транскрибируемые в пределах хроматина.
Более плотная компактизация ДНК на хроматидном и хромосомном уровне достигается за счет дальнейшей компактизации хромомеров и наблюдается в делящихся клетках с плотными хромосомами. Такая компактизация хромосом становится видна даже при использовании светового микроскопа в виде отдельных образований. Если клетка не делится, то хромосомы деспирализуются и между ними не видно границ. Диффузный материал таких хромосом называется хроматином.
Конденсация хромосом
Структура хромосом. В интерфазе хроматин обычно выявляется по периферии ядра растительной клетки или в виде сетчатых тяжей в его внутреннем пространстве. В некоторые периоды он может терять свою компактность, разрыхляться, деконден-сироваться, становясь диффузным. В интерфазном ядре при неполном разрыхлении хромосом видны участки конденсированного хроматина. Степень уменьшения плотности хроматина в интерфазе отражает функциональное состояние этой структуры. Максимальная его конденсация приводит к формированию компактных образований, получивших название митотических хромосом. Вначале они относительно инертны и отличаются от хроматина прежде всего плотностью упаковки составляющих их элементов.
В профазе в начале происходит конденсация и спирализация (скручивание) хромосом, в результате чего они становятся видимыми при микроскопии окрашенных препаратов. Увеличивается диаметр каждого завитка. Ядерная мембрана растворяется под действием ферментов, ядрышко исчезает. Центросома делится на две центриоли, после чего последние расходятся к полюсам клетки. Отмечается также фосфорилирование отдельных клеточных белков. Затем между полюсами начинает формироваться ахромати-новая фигура, похожая на веретено. Оно состоит из белка и РНК. К концу этой фазы ахроматиновая фигура вытягивается вдоль клетки, становясь веретеном. Структурно веретено представляет собой двухполюсную структуру, построенную из микротрубочек и различных белков. Хроматиды (сестринские хроматиды) удерживаются вместе центромерой. Длительность профазы составляет примерно 30-60 минут.
На рисунке 38 представлена схема цикла конденсации хромосом.
На стадии пахинемы, которая является наиболее долгой во времени стадией в мейотической профазе, происходит конденсация бивалентов и разделение каждой хроматиды надвое, в результате чего каждый бивалент представляет собой сложную спиральную структуру, состоящую из четырех сестринских хроматид (тетрад). В конце этой стадии начинается разделение спаренных хромосом-бивалентов. Теперь гомологичные хромосомы могут наблюдаться рядом. Поэтому в некоторых препаратах можно видеть четыре хромосомы, которые образуются в результате дупликации каждого гомолога, формирующего сестринские хроматиды. На этой стадии происходят обмены между гомологами и формирование хиазм.
Амитозом называется деление клетки, находящейся в состоянии интерфазы. К амитозу иногда относят все случаи немитотического деления клетки (рис. 66). При этом не происходит конденсации хромосом, распада ядерной оболочки и образования веретена деления; амитоз осуществляется при вытягивании ядра и его последующем делении на две части. Еще более неупорядоченное дробление ядра на два или более неидентичных комка получило название фрагментации; оно, безусловно, носит патологический характер. Однако между амитозом и фрагментацией резкой и принципиальной границы провести нельзя.
| Схема митотического цикла. После митоза (М) дочерние клетки вступают в фазу к концу которой происходит подготовка к синтезу ДИК. Синтез ДНК и гистонов происходит в следующей Б-фазе; в это же время происходит репликация хромосом, в результате чего содержание ДНК в клетке удваивается. Затем клетка вступает в фазу С2, представляющую собой период подготовки к митозу, включающий конденсацию хромосом. |  |
Процесс конденсации
Конденсация начинается в прометафазе, одной из стадий митоза и мейоза. Именно в этот момент, когда клеточные волокна протягиваются между пояском и полюсами, хромосомы начинают сжиматься и уплотняться. В процессе конденсации каждая хромосома становится более короткой и толстой. Это позволяет хромосомам способствовать точному разделению на две новые клетки.
Процесс конденсации сопровождается взаимодействием белков-конденсинов с ДНК. Белки-конденсины являются основными игроками в упаковке ДНК в компактные хромосомы. Они связываются с ДНК и образуют петли, а затем сворачивают их во вторичные структуры, что приводит к образованию видимых хромосомных структур.
Конденсация хромосом также регулируется хромосомными территориями. Хромосомы имеют свои зоны, где они сосредоточены в ядре клетки. Эти территории создают условия для правильной конденсации хромосом и обеспечивают правильное разделение генетической информации при делении клетки.
Таким образом, процесс конденсации играет важную роль в поддержании структуры и интегритета генома клетки
Он обеспечивает правильное упаковывание и разделение хромосом, что важно для передачи генетической информации в следующее поколение клеток
Сжатие хроматина
Сжатие хроматина происходит с помощью различных белковых комплексов, которые активно взаимодействуют с ДНК и другими белками. Один из ключевых игроков в этом процессе — белок конденсина. Конденсин является многомерным комплексом, состоящим из нескольких субединиц, и выполняет роль «прочных» связей между различными кусочками ДНК.
При сжатии хроматина белки-конденсины связываются с ДНК, образуя петли. Благодаря этому происходит формирование хромосомных структур. Такие петли сжимают нитевидное состояние хроматина, делая его более компактным и уплотненным. Это позволяет хромосомам занимать значительно меньше пространства и легко транспортироваться внутри ядра клетки.
Сжатие хроматина имеет важное значение для сохранения структуры и функций хромосом. Оно помогает защитить ДНК от повреждений и обеспечивает осуществление генальных процессов, таких как копирование, транскрипция и репарация ДНК
Кроме того, сжатие хроматина участвует в регуляции выражения генов, помогая активировать или заглушить определенные гены в разных клеточных типах и условиях.
Исследование механизмов сжатия хроматина продолжается, и по-прежнему остается много нераскрытых вопросов. Однако, уже сейчас известно, что процесс конденсации хромосом и сжатия хроматина является сложной и тщательно регулируемой системой, обеспечивающей сохранение генетической информации и ее передачу в следующее поколение.
Образование хромосомных структур
В процессе конденсации хромосом происходит сжатие хроматина — комплекса ДНК и белков, который образует хромосомную структуру. Сжатие позволяет хромосомам занимать меньшее пространство в ядре клетки и предотвращает их случайное перемещение.
Образование хромосомных структур происходит благодаря регуляции конденсации, в которой важную роль играют специальные белки-конденсины. Эти белки связываются с хроматином и обеспечивают его уплотнение и организацию в хромосомную структуру.
Влияние хромосомных территорий также играет важную роль в образовании хромосомных структур. Хромосомные территории представляют собой области ядра клетки, которые содержат группы генов и ассоциированы с определенными хромосомами. Расположение хромосомных территорий в ядре клетки влияет на организацию хромосомных структур и их взаимодействие с другими структурами в ядре.
| Функции конденсации хромосом: |
|---|
| Уплотнение генома в ядре клетки |
| Правильное размещение генов на хромосомах |
| Предотвращение случайного перемещения хромосом |
Таким образом, образование хромосомных структур играет важную роль в организации генома в ядре клетки, обеспечивая правильное размещение генов и предотвращая их случайное перемещение. Регуляция конденсации, влияние белков-конденсинов и хромосомных территорий играют ключевую роль в этом процессе.
Фаза I
Фаза I stands as a pivotal stage in meiosis, characterized by the intricate interactions of homologous chromosomes and the subsequent exchange of genetic material. This phase is further subdivided into five distinct stages, each with its unique processes and characteristics.







1. Лептотена:
- Marking the onset of Фаза I, Leptotene witnesses the condensation of replicated chromosomes.
- Эти хромосомы, теперь более компактные и различимые, напоминают нити, украшенные бусинками, называемыми хромомерами.
- Любопытно, что на этой фазе каждая сестринская хроматида прикрепляется к ядерной оболочке.
2. Зиготена (или Зигонема):
- Важная фаза, зиготена, характеризуется тесной ассоциацией гомологичных хромосом, образующих пары в процессе, называемом синапсом.
- Эти парные хромосомы, или тетрады, состоят из четырех хроматид.
- Синаптонемный комплекс, структура, похожая на застежку-молнию, образованная спиральными хроматидами, играет решающую роль в облегчении синапса, гарантируя, что хромосомы остаются выровненными.
3. Пахитена:
- Эта стадия отмечена генетическим кроссинговером между несестринскими хроматидами гомологичных хромосом, приводящим к образованию хиазм.
- Синаптонемный комплекс, выполнив свою роль в синапсе, теперь обеспечивает обмен генетическим материалом, внося вариации путем обмена материнскими и отцовскими генетическими элементами.
- Несмотря на разделение сестринских хроматид, гомологичные хромосомы остаются связанными, образуя плотную структуру, известную как синаптонемный комплекс.
4. Диплотена:
- По мере распада синаптонемного комплекса гомологичные пары хромосом остаются соединенными в хиазмах.
- Отталкивание между плечами хромосом заставляет их расходиться, однако хиазмы удерживают их вместе.
- На этой фазе происходит явление терминализации, когда хиазмы мигрируют к концам хроматид, становясь видимыми под микроскопом.
5. Диакинез:
- Кульминация Фаза I, Diakinesis prepares the cell for the subsequent metaphase.
- Хромосомы, подвергшиеся дальнейшей конденсации, под микроскопом различимы как тетрады или биваленты.
- По завершении терминализации хиазмы располагаются на концах хроматид.
- Одновременно ядрышко и ядерная оболочка распадаются, освобождая центриоли и способствуя образованию митотического веретена.
Влияние окружающей среды на конденсацию хромосом
Конденсация хромосом является важным этапом клеточного деления и интенсивно исследуется научным сообществом. Однако сложность процесса и точные механизмы его регуляции до конца не изучены. Некоторые исследования позволяют сделать выводы о том, что окружающая среда может оказывать влияние на конденсацию хромосом.
Например, известно, что изменение температуры может влиять на скорость конденсации хромосом. Понижение температуры может замедлять процесс, а повышение – ускорять его. Однако конкретная температура, при которой это происходит неизвестна.
Другие исследования показывают, что факторы окружающей среды могут влиять на конформацию хромосом до начала клеточного деления. Например, экспозиция ксенобиотиков, таких как тяжелые металлы, может привести к изменению структуры хроматина и связанным с этим изменениям в процессе конденсации хромосом. Это может привести к ошибкам в работе генетического материала, что далеко не всегда хорошо для клеток и организмов в целом.
Таким образом, существует достаточно доказательств того, что окружающая среда может оказывать влияние на конденсацию хромосом. С учетом этого можно предположить, что влияние окружающей среды должно быть учтено в ходе медицинских процедур и воздействий на организмы, чтобы минимизировать возможные риски для генетического материала и здоровья в целом.
Важность профазы
Профаза, как начальная стадия как митоза, так и мейоза, имеет первостепенное значение в процессе деления клеток. Его значение можно понять по следующим моментам:
- Хромосомная конденсация: Во время профазы волокна хроматина конденсируются с образованием отдельных видимых хромосом. Эта конденсация имеет решающее значение, поскольку она облегчает последующее упорядоченное разделение генетического материала.
- Образование сестринских хроматид.: каждая хромосома реплицируется с образованием двух сестринских хроматид, которые необходимы для равного распределения генетического материала между дочерними клетками.
- Запуск шпиндельного аппарата: Центросомы, присутствующие в животных клетках, начинают двигаться к противоположным полюсам клетки, инициируя формирование веретенообразного аппарата. Эта структура жизненно важна для выравнивания и разделения хромосом во время деления клеток.
- Разрушение ядерной оболочки: Распад ядерной оболочки гарантирует, что волокна веретена могут получить доступ к хромосомам и взаимодействовать с ними, подготавливая их к выравниванию в метафазной пластинке.
- Homologous Pairing in Мейоз: В частности, в профазе I мейоза гомологичные хромосомы объединяются в пары, что обеспечивает генетическую рекомбинацию. Этот процесс увеличивает генетическое разнообразие, необходимое для эволюции и адаптации.
- Пересекая: Еще в профазе I мейоза сегменты несестринских хроматид могут меняться местами. Этот кроссинговер приводит к генетической изменчивости, гарантируя, что потомство будет иметь уникальную комбинацию генов.
- Активация контрольной точки: Профаза также служит точкой, где клетка проверяет целостность своего генетического материала. Если обнаружено какое-либо повреждение, клетка может остановить процесс, устранить повреждение или, в крайних случаях, подвергнуться запрограммированной гибели клетки.
- Энергосбережение: Конденсируя хромосомы и разрушая определенные структуры, такие как ядерная оболочка, клетка сохраняет энергию, которую можно перенаправить на более поздние стадии клеточного деления.
- Профилактика генетических ошибок: Процессы, происходящие во время профазы, особенно контрольные точки, обеспечивают минимизацию генетических ошибок. Это имеет решающее значение для правильного функционирования организма и предотвращения таких заболеваний, как рак.
- Установление полярности: Движение центросом к противоположным полюсам устанавливает полярность, необходимую для направленного воздействия на хромосомы на последующих стадиях клеточного деления.
Основные типы компактизации хромосом
Средства электронной микрофотографии позволяют рассмотреть растительную клетку очень подробно. Так, посредством этого прибора стало возможным рассмотреть состав клетки: в ней есть темные и густые сосредоточения гетерохроматина, а вот более светлые части являются эухроматином. В процессе синтеза рибонуклеиновой кислоты при помощи ДНК, то есть трансляции информации генов из дезоксирибонуклеиновой кислоты на рибонуклеиновую кислоту. Там же можно найти центромеры и теломеры, которые составляют хромосомную структуру. В них нет никаких генов. Эти элементы считаются неизменной частью гетерохроматина. Формирование дополнительного гетерохроматина в рамках клетки совершается без неизменной базы. Из-за этого в биологии различают такие виды компактизации нуклеопротеида, который составляет хромосому:
- Нуклеосомный тип. Представляет собой два с половиной оборота удвоенной спирали дезоксирибонуклеиновой кислоты, что обвивается вокруг корда белка.
- Нуклеомерный тип (его также называют суперспиральным). Его формирует гистоновый белок. В итоге данного типа происходит создание фибриллы хроматина. Величина в обхвате у данного образования составляет примерно 30 нанометров. Получается, что ДНК произвела компактизацию на данном этапе 40 раз.
- Хромомерный тип. Происходит объединение петельного белка друг с другом посредством всех белков хроматина, кроме гистонов. Величина в обхвате подобного образования — 300 нанометров.
- Хромонемный тип белка. Формирование данного белкового типа соотносится с тем, что хрономеры приближаются друг к другу на всей протяженности линии. В нее включается единственная большая ДНК-молекула.
- Хроматидный тип белка. Посредством сложения данного белка пару раз формируется хроматидное тело. В итоге репликации в структуре дезоксирибонуклеиновой кислоты появляется около двух хроматид.
- Хромосомный тип белка, который образуется из двух хроматид. Посредством центромеры объединяются все структурные части хромосомы. После процесса клеточного деления совершается процедура хроматидного расхождения — хроматиды проникают в различные сформированные клетки, которые считаются дочерними.
Получается, что хроматин изменяет свою структуру, но поддерживает неразрывность связи поколений в хромосомах. На этапе интерфазы можно наблюдать отделение хроматина в форме небольших комочков, которые распространяются по всей ядерной нуклеоплазме. Когда происходит изменение клетки в митозе (изначально на этапе метафазы), возникает новая хромативная форма — хромосомы. Это очень ярко окрашенные тела.
Хромосома и хроматимные комочки в нуклеоплазме — два противоположных типа структуры хроматина. Их связь формируется в митотическом ряде характерными двусторонними переходами. Подтверждают это и различные биологические исследования, эксперименты, которые получаются в итоге использования технологий электронного микроскопа. В базе структуры метафазы и интерфазы находится простейшая белковая структура в виде нити.
Неравноценный уровень хромосомной компактизации обладает огромным смыслом по своему значению. Существует два типа гетерохроматина: обязательный, который присутствует всегда, и дополнительный. Значение всех видов гетерохроматина заключается в том, чтобы сохранять цельную ядерную структуру и крепить хроматиновые части к оболочке ядра. К тому же, посредством гетерохроматина происходит управление деятельностью многих структур клеток.
Что такое компактизация ДНК: система и фазы
Компактизация дезоксирибонуклеиновой кислоты — процедура укладки дезоксирибонуклеиновой кислоты в хромосому.
Хромосомы формируются на базе двух половин, которые составляют нижнее и верхнее плечо хромосомы. По своей сути хромосома является двумя хроматидами, которые соединяются посредством центромера. Приблизительная продолжительность структуры ДНК внутри человеческих клеток — 2 метра. В диаметре ядерная клетка достигает 7 микрометров. Если учитывать, что все хромосомы имеют свою ДНК-молекулу, то компактизация дезоксирибонуклеиновой кислоты происходит в шесть тысяч раз. Этот процесс происходит в несколько этапов:
- Первый уровень считается нуклеосомным. В случае, когда на хроматин влияет нуклеаз, то дезоксирибонуклеиновая кислота разрушается и будет сформирована из структур, которые постоянно повторяются. Подобные структуры представляют собой восемь белковых молекул. Эти молекулы называются гистоны и они вьются вокруг молекулы, которая имеет восемь мономерных звеньев и входит в состав ДНК. Данная структура носит название нуклеосома. Гистоны в нуклеосомы формируют основу из белка, на поверхности ее помещаются два витка ДНК. В то же время между нуклеосомами сохраняется линкер. Линкером называют часть, которая объединяет две нуклеосомы, которые находятся рядом. В научной среде этот уровень компактизации дезоксирибонуклеиновой кислоты именуют бусинами на ниточке. ДНК представляет собой нитку, а бусинами являются нуклеосомы. По диаметру нуклеосомы достигают примерно 11 нанометров. Данный уровень является структурным и формирует частоту и насыщенность упаковки ДНК.
- Второй уровень обладает формой фибриллы, компактизация составляет примерно 30 нанометров. Фибриллы находятся в митотических хромосомах, они достигают диаметра 30 нанометров. Тем не менее, может наблюдаться соленоидный вид нуклеосом, напоминающий по форме нить с несколькими витками и шагом спирали в 10 нанометров. На витках создаются фибриллы с основной полостью. У ядерного хроматина фибриллы будут достигать диаметра в 25 нанометров. Данный уровень формирует уплотнение молекул ДНК в сорок раз.
- Третий уровень — петлевые структуры, которые носят название хромомерных. Данный уровень представляет собой систему высочайшего ранга. В рамках подобных уровней особенные белковые структуры сочетаются с дезоксирибонуклеиновой кислотой. В результате формируются массивные петли. В нескольких точках наблюдаются участки сгущенного хроматина, образования в виде розеток, которые состоят из петелек с 30 нанометровыми фибриллами. Они все соединяются в сгущенном центре.
Усредненная величина розеток обычно не больше 150 нанометров. Хромомерами называют фибрилловые розетки хроматина. Все хромомеры по своей структуре представляют собой некоторое количество петель, что соединяются в центре. Хромомеры соединяются между собой и с другими частями хроматина нуклеосом. Это позволяет провести структурную компактизацию хроматина, формирует единицы хромосом, которые несут функции. Если быть точнее, то формируют гены и репликоны, которые транскрибируются в рамках хроматина.
Более густой процесс компактизации ДНК происходит на уровне хромосом и хроматид. Это возможно становится из-за того, что потом происходит компактизация хромомеров. Этот процесс можно наблюдать в клетках, которые совершают деление в кретках с густыми хромосомами. Подобную компактизацию хромосом можно увидеть при помощи микроскопа в качестве сепарированных структур. В случае, когда клетка не совершает деление, то хромосомы перестают составлять спираль и между хромосомами нельзя увидеть никаких границ. Хроматином называют рабочую структуру подобных хромосомных структур. Белок хромосомы формируется при помощи двух хроматид. Они объединены центромерой, в процессе клеточного деления хроматиды разъединяются, перемещаясь в различные созданные клетки.
Механизмы деспирализации
Деспирализация хромосом является важным процессом в клеточном развитии и происходит в несколько этапов.
- Распаковка хромосомы
Первый этап деспирализации заключается в распаковке плотно спирализованной хромосомы. Хромосома состоит из ДНК, которая образует спиральную структуру вокруг белковых комплексов, называемых гистонами. При распаковке хромосомы гистоны разделяются, позволяя ДНК распрямиться и доступным для репликации и транскрипции.
Развитие хроматина
После распаковки, хромосома становится состоянием называется «хроматиной». Хроматин представляет собой структуру, в которой ДНК связана с различными белками и ферментами. Хроматин имеет несколько степеней укорочения и спиральности, что позволяет контролировать доступность генов для экспрессии.
Свертывание хроматина
Свертывание хроматина происходит в разных масштабах. На самом мелком уровне, фрагменты ДНК сворачиваются в миниколчанки, состоящие из около 8 нуклеосом. Около 6-7 миниколчанок затем свертываются в более плотные структуры, называемые соленоидами. Наиболее тугое сворачивание достигается на уровне поперечного адзективного сворачивания.
Интеракция с другими хромосомами
Хромосомы взаимодействуют друг с другом, чтобы обеспечить более компактное укладывание в ядре клетки. Они образуют территорию, где каждая хромосома находится в своем определенном месте. Взаимодействие между хромосомами может быть регулировано специфическими белками и ферментами.
Механизмы деспирализации и укладки хромосом имеют важное значение для правильного функционирования клеток, поскольку они позволяют доступ генетической информации к клеточным процессам, таким как репликация ДНК и экспрессия генов
Хроматин и его структура
Итак, мы установили, что нуклеиновые кислоты содержатся в веществе хромосом — структурных единицах наследственности. Их хроматин под электронным микроскопом имеет вид гранул или нитевидных образований. Он содержит, кроме ДНК, еще и молекулы РНК, а также белки, проявляющие основные свойства и названные гистонами. Все вышеперечисленные нуклеосом. Они содержатся в хромосомах ядра и называются фибриллами (нити-соленоиды). Подводя итог всему вышесказанному, определим, что такое хроматин. Это комплексное соединение и специальных белков — гистонов. На них, как на катушки, накручиваются двухцепочечные молекулы ДНК, образуя нуклеосомы.
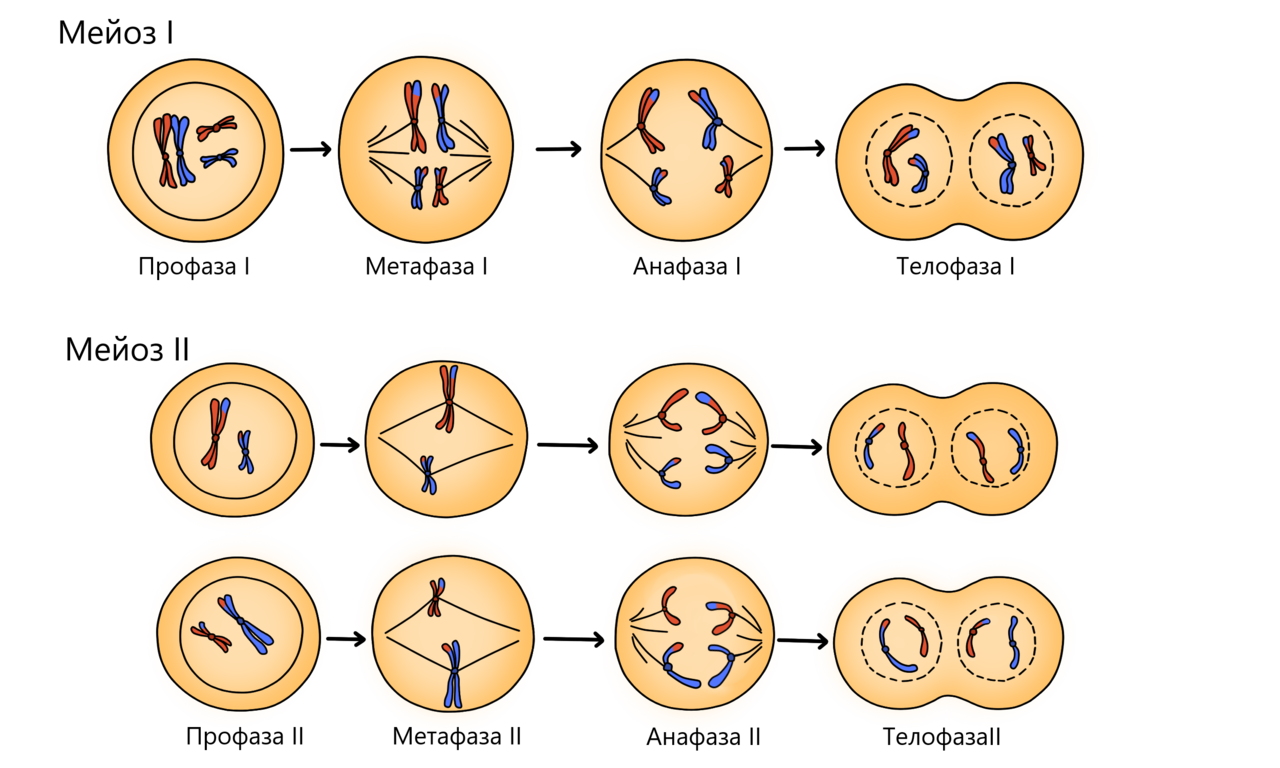
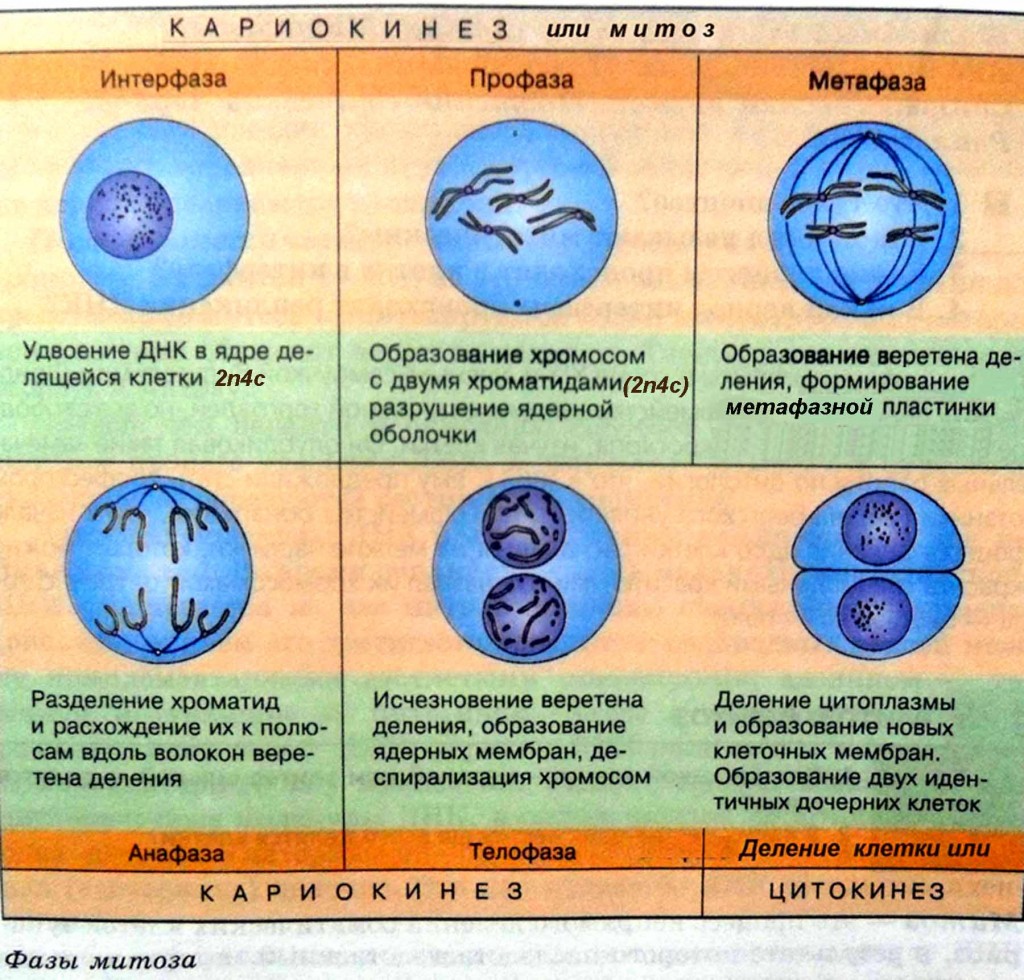


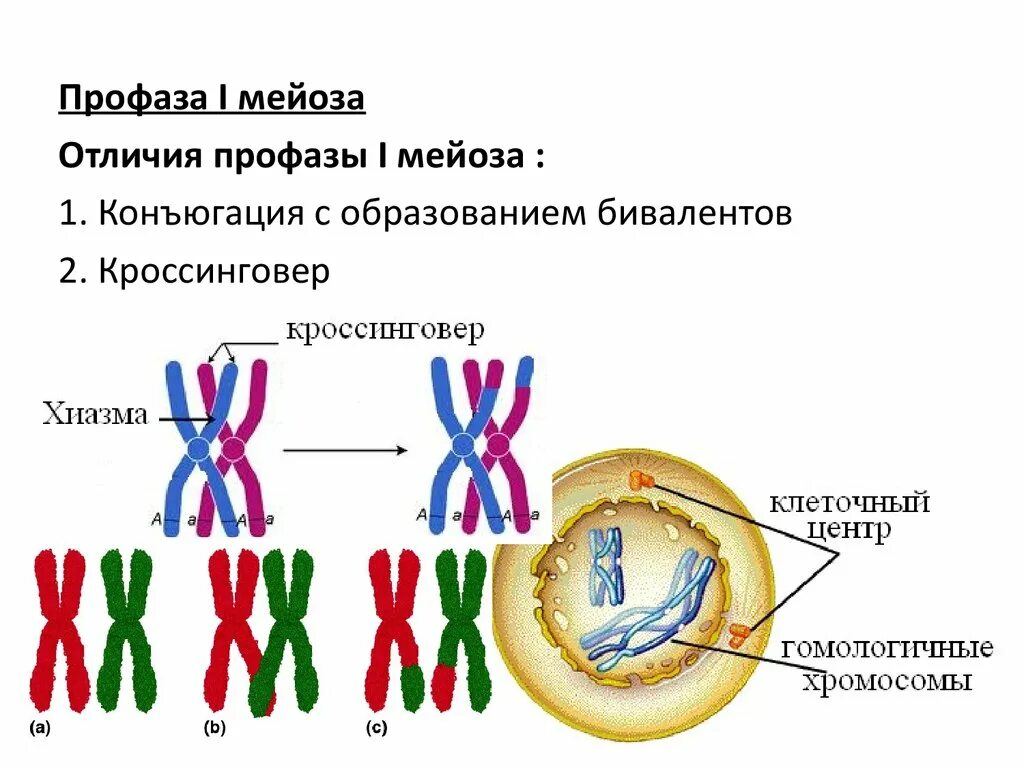


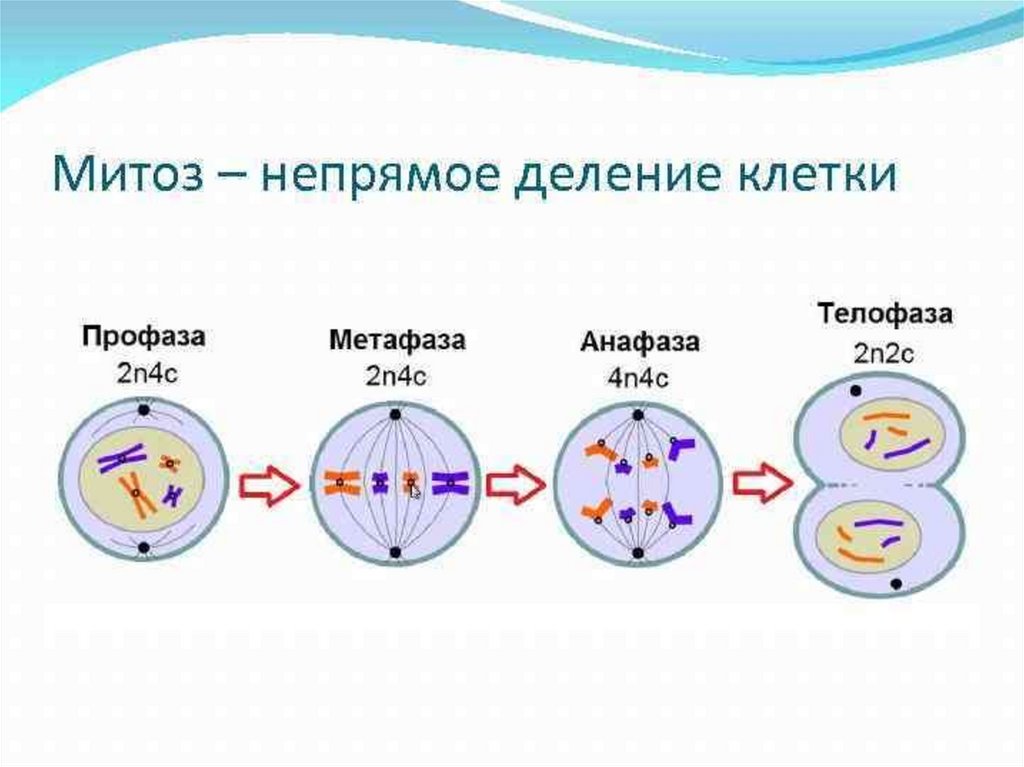
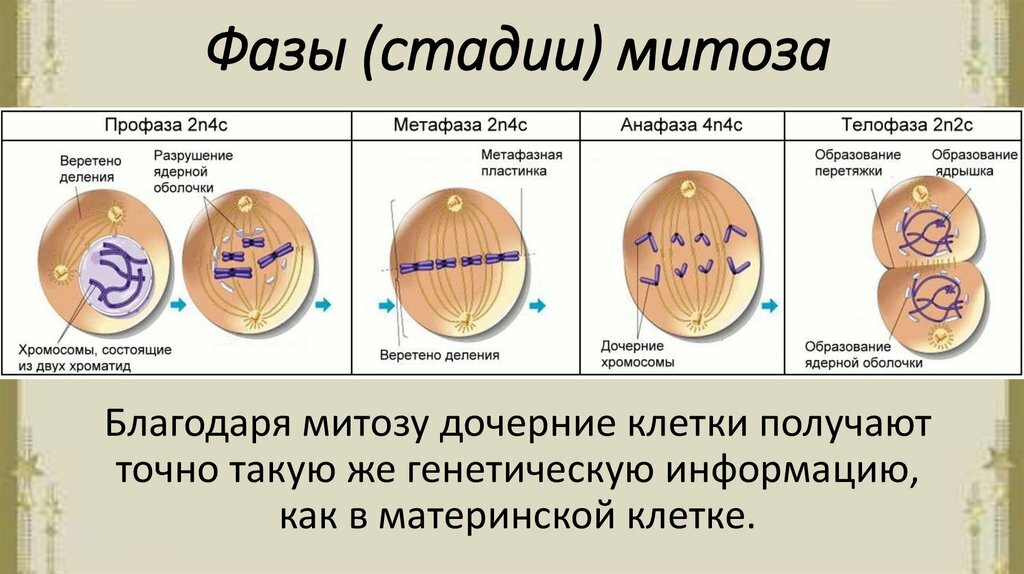
Стадии митоза
Как уже отмечалось ранее, митоз делится на 4 стадии, но иногда их может быть и больше. Ниже представлены основные из них.
Таблица. Описание основных фаз митоза.
| Название фазы, фото | Описание |
|---|---|
| Во время профазы происходит спирализация хромосом, в результате чего они принимают скрученную форму (она более компактная). Останавливаются все синтетические процессы в клетке организма, поэтому рибосомы уже не вырабатываются. | |
| Многие специалисты не выделяют прометафазу как отдельную фазу митоза. Нередко все процессы, которые в ней происходят, относят к профазе. В этот период цитоплазма окутывает хромосомы, которые свободно перемещаются по клетке до определенного момента. | |
| Следующая фаза митоза, которая сопровождается распределением на экваториальной плоскости конденсированных хромосом. В этот период происходит обновление микротрубочек на постоянной основе. При метафазе хромосомы расположены так, что их кинетохоры находятся в ином направлении, то есть направлены к противоположным полюсам. | |
| Данная фаза митоза сопровождается отделением хроматид каждой из хромосом друг от друга. Нарастание микротрубочек прекращается, они теперь начинают разбираться. Анафаза длится недолго, но за этот промежуток времени клетки успевают разойтись ближе к разным полюсам в примерно равном количестве. | |
| Это последняя стадия, на протяжении которой начинается деконденсация хромосом. Эукариотические клетки завершают свое деление, а вокруг каждого набора хромосом человека образовывается специальная оболочка. При сокращении сократительного кольца происходит разделение цитоплазмы (в медицине этот процесс называется цитотомией). |
Когда впервые в истории появилось упоминание о ДНК
Иоганнес Фридрих Фишер – врач и биолог-исследователь родом из Швейцарии, стал первым в мире ученым, выделившим нуклеиновую кислоту. Открытие случилось в 1869 году, когда он занимался изучением животных клеток, а именно лейкоцитов, которых много содержалось в гное. Совершенно случайно молодой ученый заметил, что при отмывании лейкоцитов с гнойных повязок от них остается загадочное соединение. Под микроскопом Иоганн обнаружил, что оно содержится в ядрах клеток. Это соединение Мишер назвал нуклеином, а в процессе изучения его свойств переименовал в нуклеиновую кислоту, из-за наличия свойств, как у кислот.
Роль и функции только открытой нуклеиновой кислоты были неизвестны. Однако многие ученые того времени уже высказывали свои теории и предположения о существовании механизмов наследования.
Нынешние взгляды на состав молекулы ДНК ассоциируются у людей с именами английских ученых Джорджа Уотсона и Фрэнсиса Крика, которые открыли структуру данной молекулы в 1953 году. За несколько лет до этого, в тридцатые годы, ученые из советского союза А.Н. Белозерский и А.Р. Кезеля доказали наличие ДНК в клетках во всех живых организмах, тем самым они опровергли теорию о том, что молекула ДНК находится только в клетках животных, а в клетках растений присутствует только РНК. Лишь спустя несколько лет, в 1944 году, группой освальдских ученых было установлено, что молекула ДНК является механизмом сохранения наследственной информации клетки. Таким образом, благодаря совместным усилиям и трудам исследователей человечество познало тайну процесса эволюции и его основных принципов.
Наследственность — основное свойство живой материи
К главным процессам, характеризующим организмы, живущие на Земле, относятся дыхание, питание, рост, выделение и размножение. Последняя функция является наиболее значимой для сохранения жизни на нашей планете. Как не вспомнить, что первой заповедью, данной Богом Адаму и Еве была следующая: «Плодитесь и размножайтесь». На уровне клетки генеративная функция выполняется нуклеиновыми кислотами (составляющее вещество хромосом). Эти структуры будут рассмотрены нами в дальнейшем.
Добавим также, что сохранение и передача наследственной информации потомкам осуществляется по единому механизму, который совершенно не зависит от уровня организации особи, то есть и для вируса, и для бактерий, и для человека он универсален.
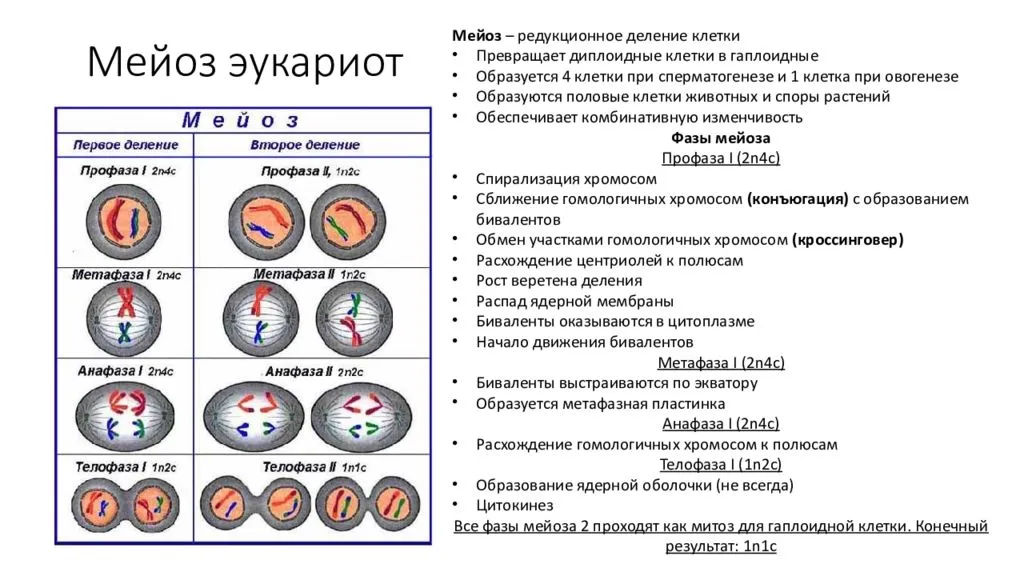
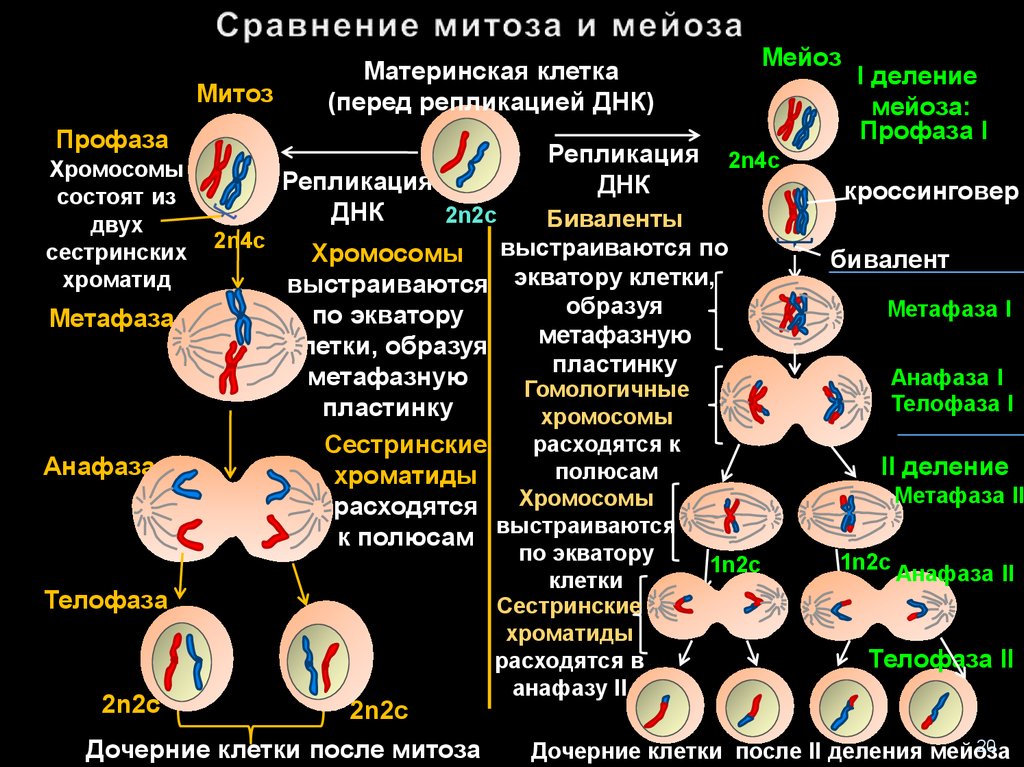
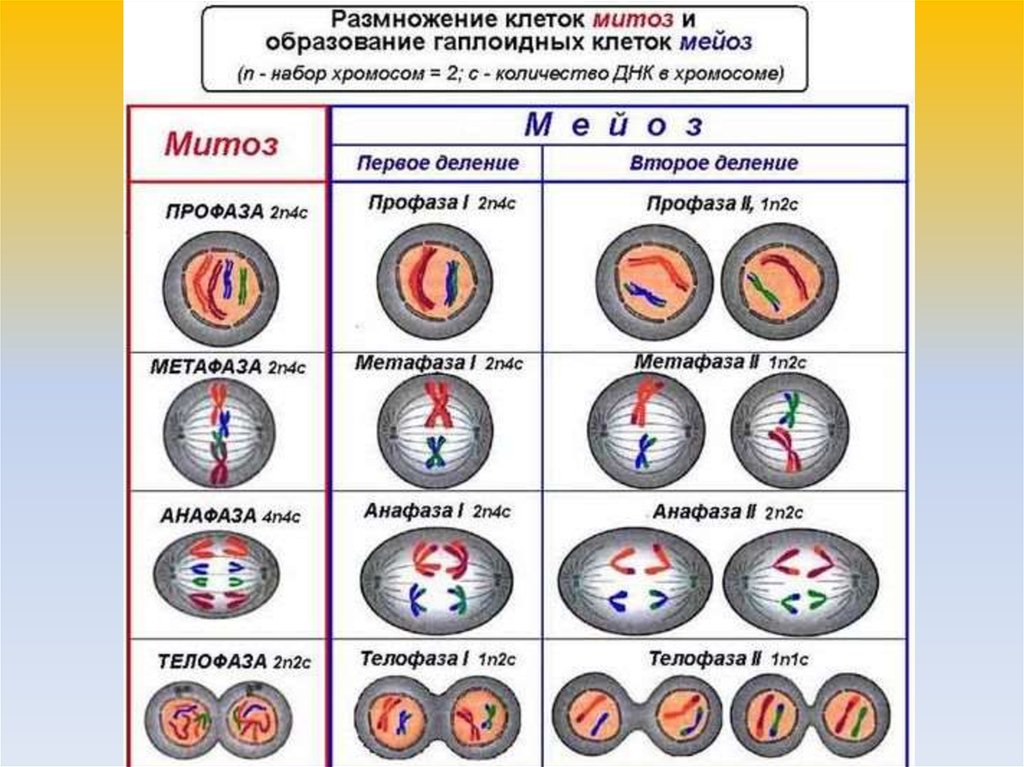
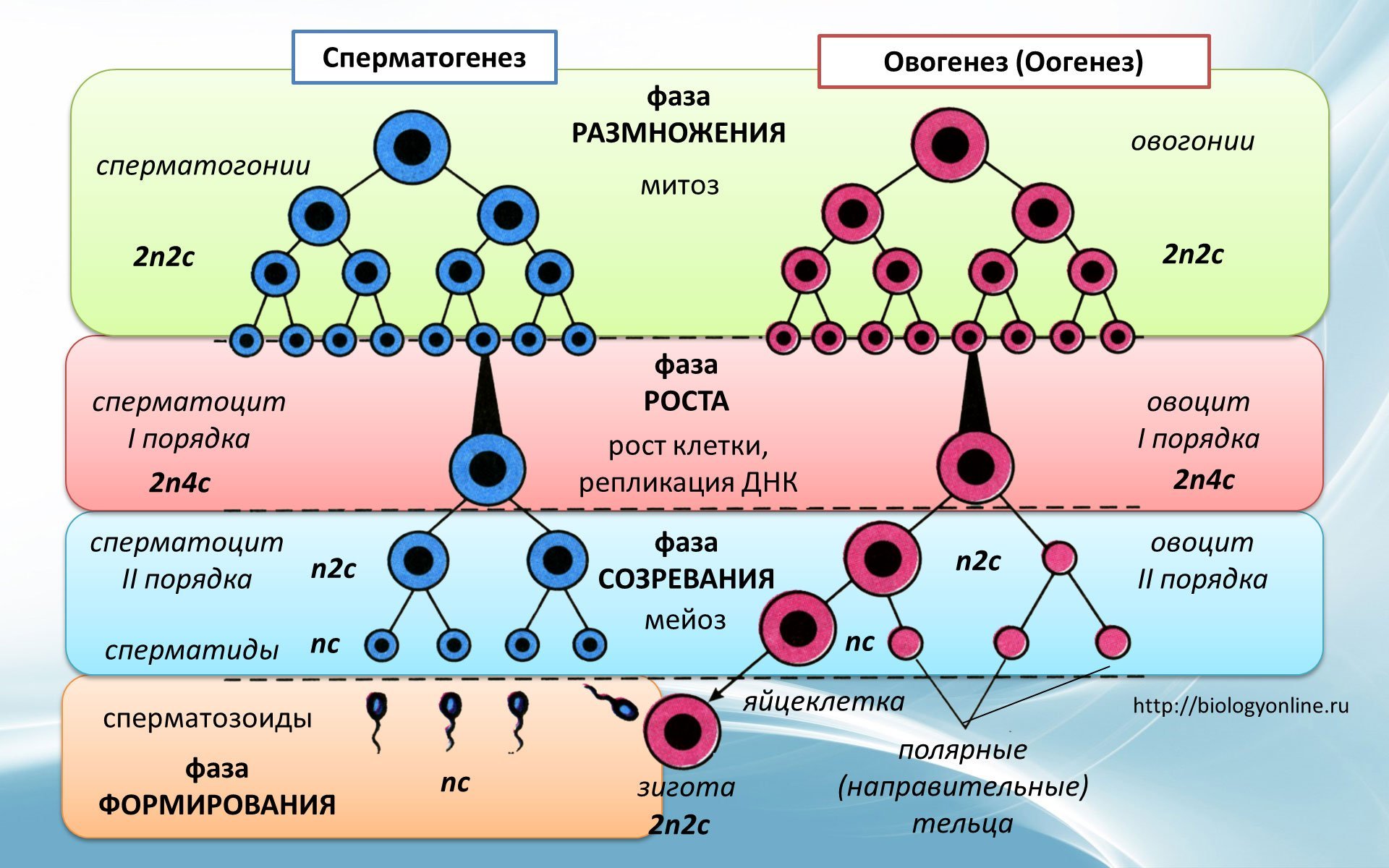
Редукционный этап или первое деление мейоза
Его суть — изменение числа хромосом внутри клетки. То есть из одной диплоидной (2n4c) клетки получаем две гаплоидных (1n2c). Так стоп, откуда 4c? До этого же было 2n2c. Ах да… Сейчас разберемся.
Интерфаза
Перед вступлением в мейоз клетка проходит через интерфазу. Ей нужно подготовиться к делению — запасти энергетических субстратов (АТФ), синтезировать необходимые белки и удвоить количество молекул ДНК. Еще в интерфазу происходит удваивание центромер.

Рисунок. Хромосомный набор в интерфазу
В интерфазу произошла репликация ДНК — образовалась идентичная цепь. Но эти две цепи, или хроматиды, связаны между собой при помощи центромеры, значит количество хромосом такое же. Итого набор — 2n4c
Ну вы ведь понимаете, что таким образом реплицируются все 46 хромосом. Просто удобнее показать на паре. Помните, что все 23 пар вступают в мейоз, а не только одна. После репликации начинается собственно мейоз, а именно его первая фаза:
Профаза мейоза I
В отличие от митоза состоит из пяти стадий: лептотена, зиготена, пахитена диплотена и диакинез. Она более длительная и здесь протекают важные процессы: конъюгация и кроссинговер. Еще в эту фазу растворяется ядерная оболочка и формируется веретено деления, подробнее об этом ниже.
Лептотена
Какая основная задача у клетки? Правильно, передать генетический материал своим потомкам. Поэтому она начинает упаковывать молекулы ДНК как можно плотнее, она собирает чемодан, ведь не хочет ничего не потерять в пути. Этот процесс называется спирализация или конденсация хромосомы. Клетка так старается, что невидимые раньше в микроскоп хромосомы становятся видимыми. Они похожи на длинные и тонкие нити.
Зиготена
Здесь происходит конъюгация хромосом — их сближение с образованием бивалентов. Связь обеспечивает синаптонемальный комплекс — он удерживает гомологичные хромосомы рядом это необходимо для запуска кроссинговера на следующем этапе.

Схема. Образование бивалентов.
Связи между хромосомами могут иметь разный вид, но они должны быть. Если в клетке останутся хромосомы, которые не сблизились, то она запускает апотоз и погибает. Клетка — с заботой о будущих поколениях!
Пахитена
Начинается с еще большей конденсации хромосом, они становятся короче и толще. Но в местах образования синаптонемальных комплексов происходит частичное раскручивание (деконденсация) хромосом.


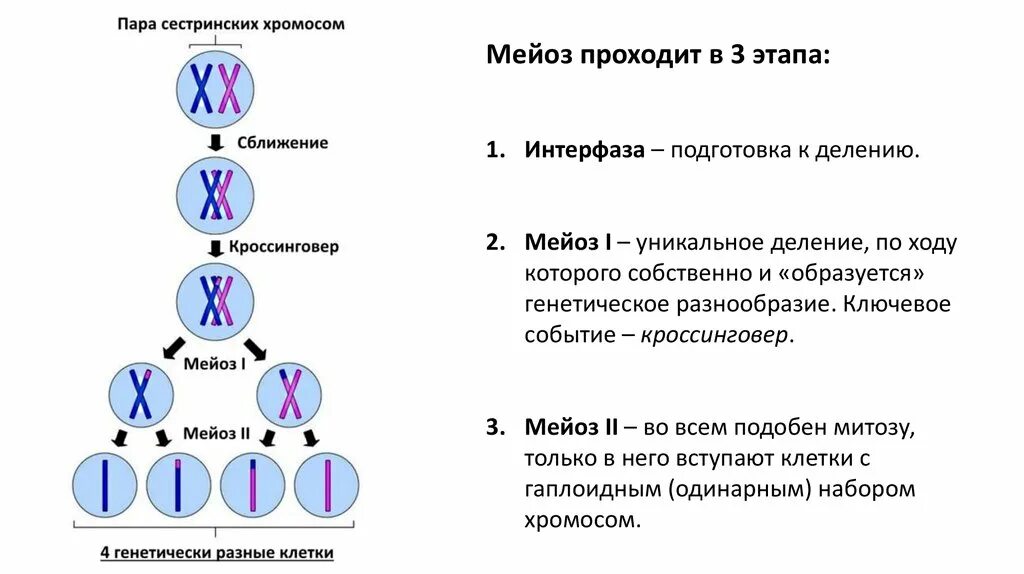



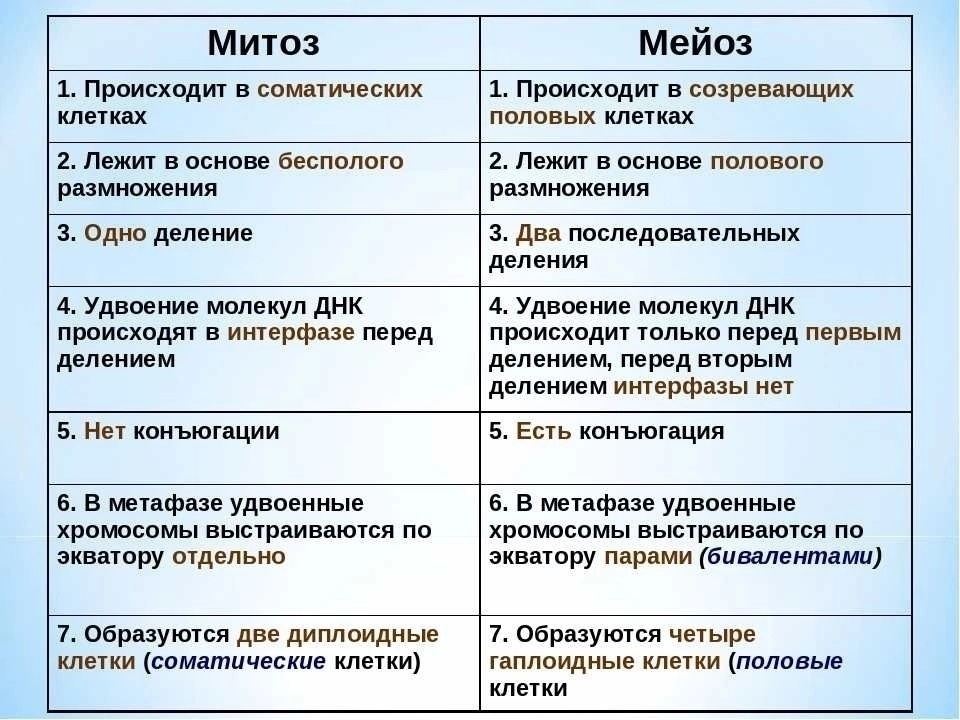
Все это для начала кроссинговера — обмена участками ДНК у гомологичных хромосом. Обмен обеспечивает перекомбинацию генетического материала. Если бы мы могли рассоединить хромосомы сразу после кроссинговера, то увидели примерно такую картину:
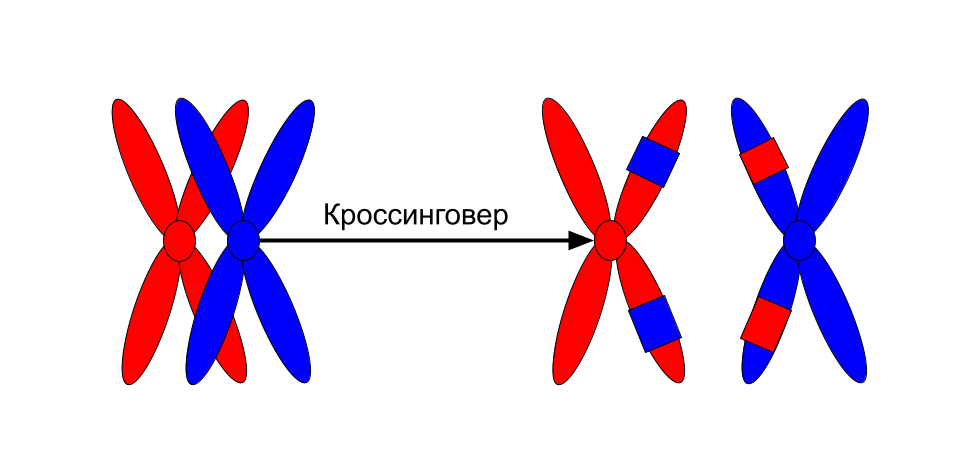
Схема. Кроссинговер.
Это лишь схематичное изображение, перекресты могут происходить в самых разных местах , что дает огромную генетическую вариабельность.
В конце пахитены мостики между хромосомами разрушаются, они начинают отдаляться друг от друга.
Диплотена
Хромосомы расходятся в области центромер, но остаются связаны между собой в местах кроссинговера — перекрестах или хиазмах. В микроскоп можно увидеть все четыре хроматиды, так сильно они упаковались (спирализовались).
Диакинез
Гомологичные хромосомы расходятся, формируется веретено деления и исчезает ядерная оболочка. Этим завершается профаза мейоза I. Вид клетки примерно такой:

Схема. Конец профазы мейоза I
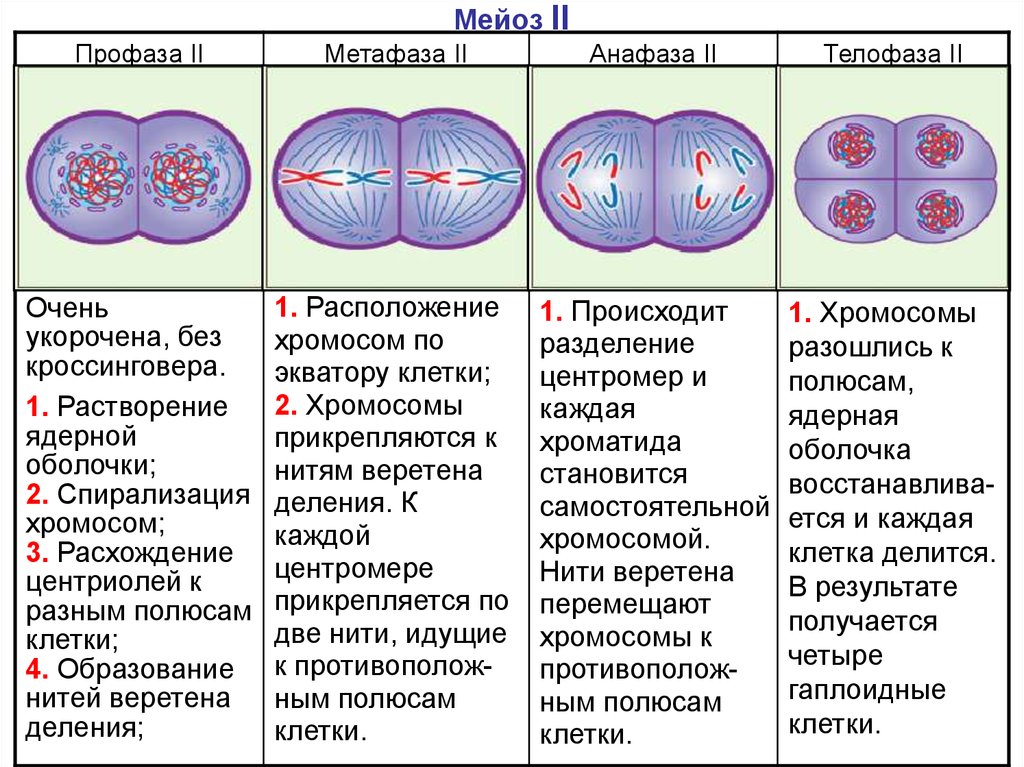
Метафаза мейоза I
В этой фазе заканчивается образование веретена деления. Нити веретена прикрепляются к центромерам и начинают притягивать хромосомы, из-за этого они располагаются на экваторе клетки.

Схема. Клетка в метафазу I
Анафаза мейоза I
Нити веретена деления продолжают тянуть хромосомы на себя — они расходятся к полюсам клетки. На полюсах клетки располагается по 23 хромосомы, но они все еще состоят из двух нитей ДНК.

Схема. Анафаза мейоза I
Телофаза мейоза I
Завершение редукционного деления. Появляется ядерная оболочка, которая окружает хромосомы. Затем возле ядер появляется перетяжка, которая делит клетку на две части. Образуются две гаплоидные клетки.
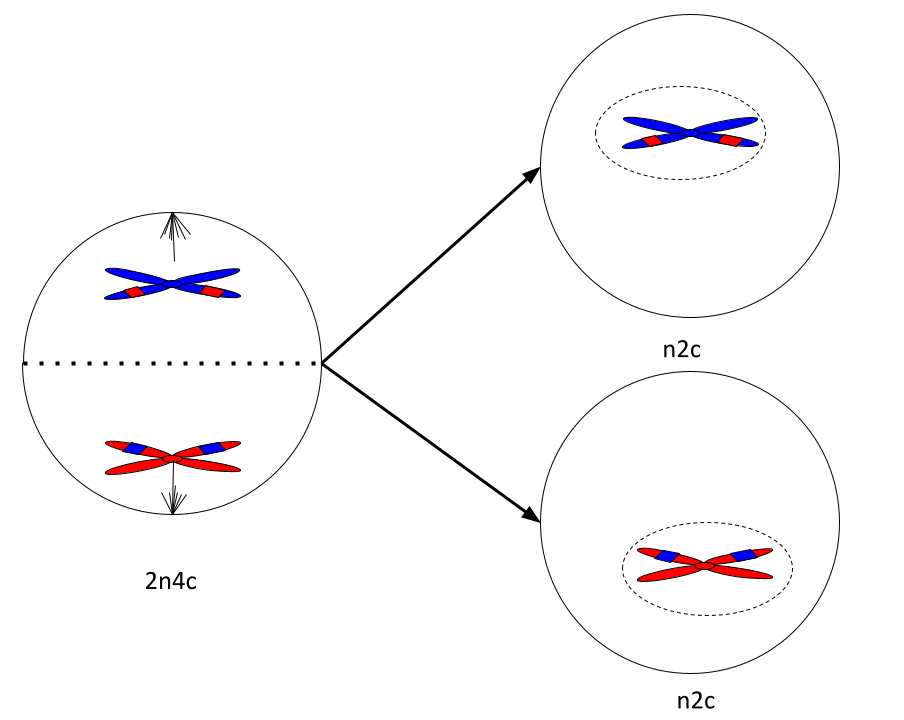
Схема. Конец первого деления мейоза